
上: ユキノシタ(ユキノシタ科)の左右相称花
右上: セイヨウアブラナ(アブラナ科)の十字相称花
右: アジサイ(ユキノシタ科|アジサイ科)の放射相称花




花の特徴によって訪れる送粉者の動きが制御される場合がある。例えば、ホウセンカやツリフネソウの花では、ハナバチが花のどこに脚を掛け、どのように動いて蜜を目指すかは、ほぼ決まっている。
送粉者の制御には、次のような要素が関わっている。
A~Cによって送粉者の動線が決められ、それに合うようにDが設定される。
訪花者の制御には、次のような利点がある。
訪花者の行動を制御する花は、横か斜めに咲き、花を正面から見たときに、明確な上下があり、制御の対象である訪花者の上下と一致している。このような花を「左右相称花」という。


花は、正面から見たときの対称面の数によって、以下の3つに分けられる。
上: ユキノシタ(ユキノシタ科)の左右相称花 右上: セイヨウアブラナ(アブラナ科)の十字相称花 右: アジサイ(ユキノシタ科|アジサイ科)の放射相称花 |
|
左右相称の程度はいろいろで、ユキノシタやホウセンカの花のように非常にはっきりと花の上下の違いがあるものを「強い左右相称」、サツキのように比較的違いが弱いものを「弱い左右相称」という。

ハナバチ媒の蜜花には、典型的な制御のしくみを持つものが多い。こういう花では、ハナバチが着地する場所や蜜を求めて体を動かす道筋が決まっている。葯と柱頭は互いに接近していて、ハナバチが訪花し、吸蜜して去るまでの一連の動きの中で体表のほぼ同じところに葯と柱頭が触れる。ハナバチが同種の別の個体を訪花すれば、高い確率で送粉に成功することになる。
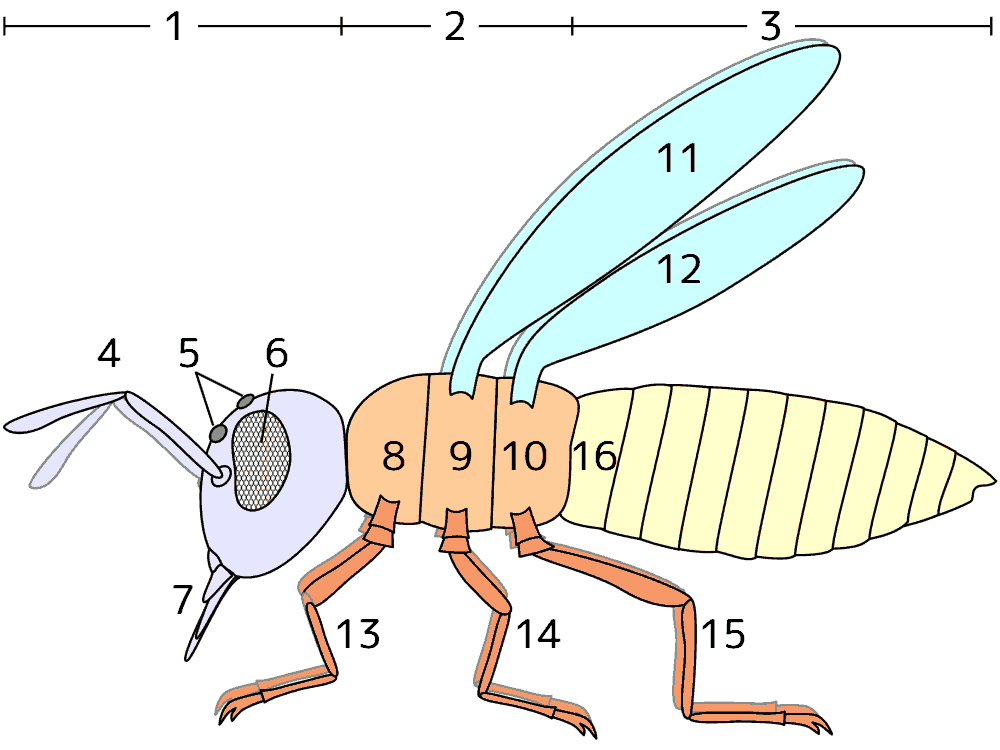 昆虫の体の模式図
昆虫の体の模式図葯・柱頭の付着部位で分けると、次の2つが多い。
ハナバチは毛に覆われた体表についた花粉を脚を使って後肢の花粉籠にかき集めるが、上のような場所には脚が届きにくい。
背面摩擦型の花の方が例が多く、ツリフネソウ属(ホウセンカの仲間)・ラン科・アヤメ科・スイカズラ科・キキョウ科・シソ科などさまざまなグループに見られる。トンネルのような送粉者の通り道の天井から柱頭や葯が下向きに突き出している形の花が多い。

 ホウセンカ(ツリフネソウ科)の花は、花被片が組み合わさってラッパのようになり、入口にちり取りのようなひらひらがついている。花の後ろ側には細長いひものような距(きょ)がついている。花を正面から見ると、入口の上から葯(開花前期)または柱頭(開花後期)が突き出していて、ホウセンカの花にもぐり込むミツバチの背中に触れる。
ホウセンカ(ツリフネソウ科)の花は、花被片が組み合わさってラッパのようになり、入口にちり取りのようなひらひらがついている。花の後ろ側には細長いひものような距(きょ)がついている。花を正面から見ると、入口の上から葯(開花前期)または柱頭(開花後期)が突き出していて、ホウセンカの花にもぐり込むミツバチの背中に触れる。ラン科の花では、ハナバチは足場として特殊化した花被片「唇弁」(リップ)[lip]上を、奥の蜜を吸うために往復する(シランのように、見かけのみで蜜を出さない種類も多い)。唇弁の真上には、雄しべと雌しべが合わさって1本の柱のようになった蕊柱―ずいちゅう―(カラム)[column]があり、蕊柱の先端にある柱頭と花粉塊がハナバチの経路に張り出している。→シラン(ラン科)の花




 シランが咲いている周辺では、花粉塊が背面に付着したハナバチがひんぱんに見られる
シランが咲いている周辺では、花粉塊が背面に付着したハナバチがひんぱんに見られるアヤメ科アヤメ属(アヤメ・ノハナショウブ・キショウブ・アイリス類など)の花は、背面摩擦型の花が3つ集まったものに相当する。→アヤメ属の花
 ノハナショウブの花にもぐり込むトラマルハナバチ
ノハナショウブの花にもぐり込むトラマルハナバチ

腹面摩擦型は、背面摩擦型と比べるとやや少なく、マメ科・ケマンソウ科・ヒメハギ科・シソ科などに例が見られる。葯と柱頭は上を向いていて、ハナバチの足場となる花弁に収納されている。ハチが乗ると花弁が押されて下がり、葯と柱頭が突き出す。
マメ科の「蝶形花」と呼ばれる花では、蜜は旗弁(上にある大きな花びら)の付け根の奥にある。ハナバチが旗弁の下の2対の花びらを足場にして潜り込むと、足場となった花びらがぐっと下がり、中に包まれている葯と柱頭が露出して腹面に触れる。
 ゲンゲ(レンゲソウ)(マメ科)の花
ゲンゲ(レンゲソウ)(マメ科)の花
 吸蜜するミツバチの腹部に、雄しべとめしべの先端が押しつけられている
吸蜜するミツバチの腹部に、雄しべとめしべの先端が押しつけられているシソ科の草本では花冠の先端が上下にぱっくりと分かれることが多く、「唇形花冠」と呼ばれる(→シソ科の唇形花)。上の方を「上唇」、下の方を「下唇」という。シソ科の唇形花では、背面摩擦型と腹面摩擦型の両者が見られる。

オドリコソウ(シソ科)では、花びらは一体化して先が曲がった筒になり、先がくちばしのように上下に割れて、上唇は傘のようになり、下唇はひれ状になっている。葯と柱頭は、傘の下に隠れている。ハナバチが下唇に着地して筒にもぐり込んで蜜を吸うときに、葯と柱頭はハチの背中に触れる。





ヤマハッカ(シソ科)では、オドリコソウと逆に、上唇がひれ状、下唇は二つ折りになっていて、葯・柱頭は下唇の中に収まっている。ハナバチが下唇に着地して筒の中に潜り込むときに、葯と柱頭がハチの腹に触れる。

ウマノスズクサ(ウマノスズクサ科)やマムシグサ(サトイモ科)は、昆虫を花(ウマノスズクサ)や花序(マムシグサ)の中に閉じこめ、動き回らせて受粉する(トラップ送粉[trap pollination])。ウマノスズクサの花は両性花で花粉をつけて昆虫を解放する。雌雄異株のマムシグサでは、雄花序は昆虫を解放するが、雌花序からは昆虫は逃げられない。
 咲き始めのウマノスズクサの花(断面)。雌しべ・雄しべがある部屋への通路には斜めの毛が生えており、逆行の妨げになる。時間が経つと柱頭が閉じて代わりに葯から花粉が出てくる。そのころになると毛がしなびて通路から出られるようになる。
咲き始めのウマノスズクサの花(断面)。雌しべ・雄しべがある部屋への通路には斜めの毛が生えており、逆行の妨げになる。時間が経つと柱頭が閉じて代わりに葯から花粉が出てくる。そのころになると毛がしなびて通路から出られるようになる。

 マムシグサの花序は深いカップ状に丸まった葉(仏炎苞)に包まれている。ハエの仲間が仏炎苞の先から入り、下に降りて小さい花が密集している空間に入ると、仏炎苞の「返し」に阻まれて上に戻ることができず、出口を探して動き回る。
マムシグサの花序は深いカップ状に丸まった葉(仏炎苞)に包まれている。ハエの仲間が仏炎苞の先から入り、下に降りて小さい花が密集している空間に入ると、仏炎苞の「返し」に阻まれて上に戻ることができず、出口を探して動き回る。| 雄花序 | 雌花序 |
  雄花の花粉に塗れたハエは仏炎苞基部の小さなすきまから脱出できる |
   仏炎苞にはすきまがなく、ハエは脱出できずに力尽きる |
花による訪花者の制御は、常に成功するわけではない。
送粉シンドロームに合わない訪花者が、選別をすり抜けたり、強行突破することもある(選別の失敗)。このような場合、制御もうまくはたらかない。また、花に適合した訪花者であっても、花の形態から予測されるような行動を取らず(制御の失敗)、訪花者≠送粉者となることがある。
 ヤマフジ(マメ科)の花弁のすきまから長い嘴を突き刺して吸蜜するセダカコガシラアブ。
ヤマフジ(マメ科)の花弁のすきまから長い嘴を突き刺して吸蜜するセダカコガシラアブ。


スイカズラ(スイカズラ科)はさまざまな昆虫の訪花を受ける(前の方で述べたように、夜間はスズメガによって送粉される)。花に潜り込んで吸蜜するコマルハナバチは、コマルハナバチは腹端の毛に花粉がついており、おそらく有効な送粉者だ。

しかし、一方では、スイカズラの花筒に孔を空けて吸蜜する(盗蜜する)コマルハナバチもいる。盗蜜をするハチは盗蜜ばかりを繰り返す傾向があり、腹端には花粉がついていない。

この例のように、同種の訪花者であっても送粉する個体と盗蜜する個体に分かれる場合がある。

逆に、盗蜜者や花に潜り込まずに花粉だけを食べる訪花昆虫がが送粉に関与することもある。だから、選別・制御に失敗しても、送粉が必ず失敗するとは限らない。

動物送粉は一筋縄ではいかない複雑な現象だ。同じ種の花であっても時期や地域によって訪花者は違うことがある。また、同じ訪花者であっても行動には個性がある。