

ノビル(ユリ科|ヒガンバナ科)はネギの仲間で、花序もネギ坊主になる。しかし、花序の一部で花の代わりにむかごができることが多い。下右のように、ほとんどむかごばかりになることもある。


前の方で述べたように、被子植物の花はシュートの先端が変形したもの、そして花被片・雄しべ・心皮(雌しべの構成要素)は葉が変形したものと考えられている。
この解釈は、ゲーテ(1749-1832)の『植物変態論』(1790)で丹念に論証され、また、裏付ける(この説を使うと説明しやすい)観察結果も、たくさん知られている。
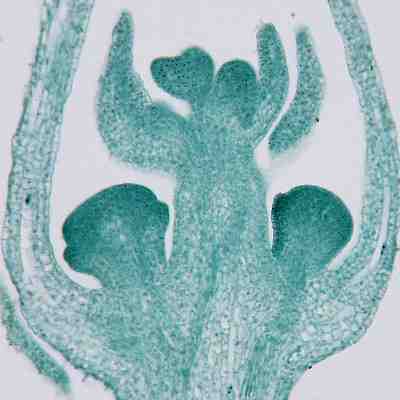 ムラサキケマン(ケシ科)の形成中の花序。花の原基が葉腋にできている。
ムラサキケマン(ケシ科)の形成中の花序。花の原基が葉腋にできている。
花は、シュートの先端、つまり、主軸の頂端または葉腋につく。ただし、花序の葉(花苞[floral bract])は普通の葉よりもずっと小さいことが多く、ない場合もある。
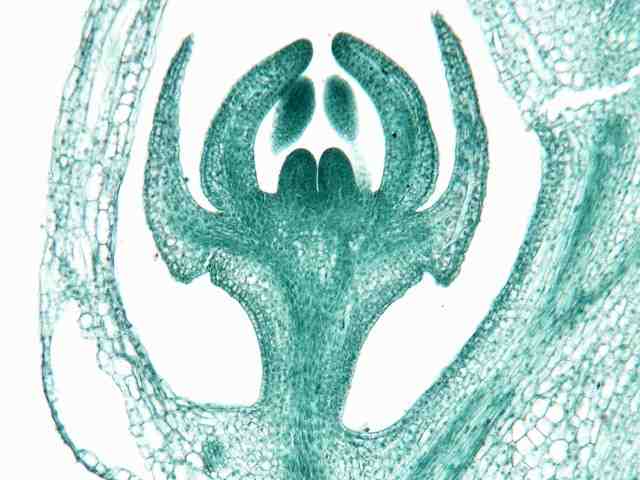 ムラサキケマン(ケシ科)の花の原基
ムラサキケマン(ケシ科)の花の原基
花の原基は、シュートの原基とよく似ている。頂端を取り巻くように花被片・雄しべ・雌しべの順に原基が形成される。
また、花の原基が環境条件によって芽に変わり、むかごとしてはたらくノビルのような例もある。
| ノビル(ユリ科|ヒガンバナ科)はネギの仲間で、花序もネギ坊主になる。しかし、花序の一部で花の代わりにむかごができることが多い。下右のように、ほとんどむかごばかりになることもある。 |
| |
維管束の通り方が、花の中心軸は茎と、萼片・花弁・雄しべ・雌しべは葉とよく似ている。ただ、これには例外もある。
現生裸子植物や化石裸子植物の胚珠錐は、分岐した多数のシュートからなる複雑なつくりを持つ。また、モクレン科のように、花の維管束が分岐したシュートに似たパターンを示すものもある。
これらのことから、花は単一のシュートではなく分岐による複数のシュートの集合体から進化した、という説が繰り返し唱えられてきたが、現在ではほとんど支持されていない。
萼片と花弁がほとんど同じような形をしている例や中間型を介して連続的な例は少なくない。また、雄しべが小さな花弁のような形をしていて、そこに葯がついているものがある。そういう場合には、雄しべと花弁の中間的な器官がしばしば見られる。
 スイレン(スイレン科)。花被片と葯を並べると、連続的に移行しているようすが分かる。いちばん外側の花被片は厚ぼったくて裏側が緑色を帯びている。花弁状の花被片は内側のものほど小さく、最外の雄しべは花弁形をしている。
スイレン(スイレン科)。花被片と葯を並べると、連続的に移行しているようすが分かる。いちばん外側の花被片は厚ぼったくて裏側が緑色を帯びている。花弁状の花被片は内側のものほど小さく、最外の雄しべは花弁形をしている。


雄しべと雌しべのあいだに中間的な器官が見られるものもある。


 ハクモクレン(モクレン科)。ドーム状に盛り上がったところ(花托)の下の方に雄しべが、上の方に雌しべがついている。雄しべは縁からクリーム色の花粉を出す。雌しべの先端は細長く伸びたピンク色の花柱となる。
ハクモクレン(モクレン科)。ドーム状に盛り上がったところ(花托)の下の方に雄しべが、上の方に雌しべがついている。雄しべは縁からクリーム色の花粉を出す。雌しべの先端は細長く伸びたピンク色の花柱となる。
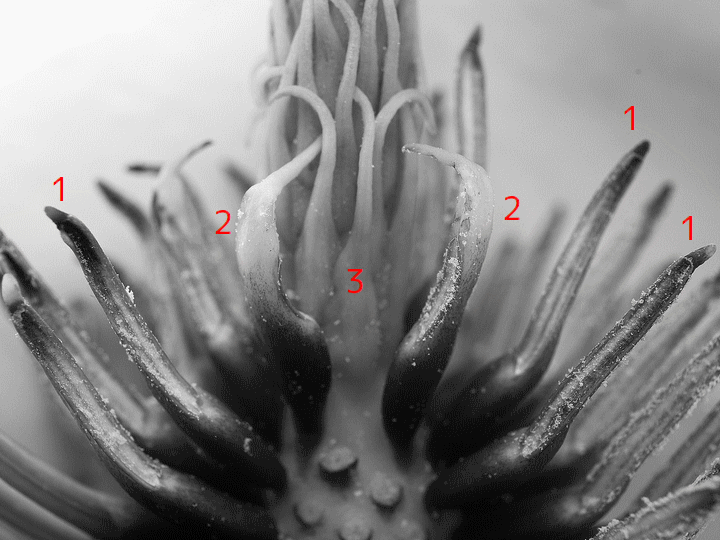
園芸では、ふつうの花では見られない特徴を持つ花が「変わり花」(奇形花)と呼ばれて古くから珍重されてきた。変わり花の中には、萼片が花弁になったり(またはその逆)、花弁が雄しべになったり(またはその逆)しているものがある。また、萼片・花弁・雄しべが葉のようになったものや、雌しべが複数の葉になっているものがある。
 八重咲きのツバキ。雄しべが花弁に変わっているため、花弁がもともとの数(約5)より多い。ふつうの(八重咲きでない)ツバキの画像はここ。
八重咲きのツバキ。雄しべが花弁に変わっているため、花弁がもともとの数(約5)より多い。ふつうの(八重咲きでない)ツバキの画像はここ。
例えば、変わり花の代表格の「八重咲き」[double flower]は、雄しべ(ときに雌しべも)が花弁になった個体だ。有性生殖の能力が低いために野生で増え続けるのは難しいと考えられ、実際にも、野外で自生している植物で八重咲きの花はごくまれにしか見つからない。人為的に保存し、挿し木などで増殖することで多数の八重咲品が存在している。





器官がまるまる別の器官に置き換わる現象をホメオシス[homeosis]という。ホメオシスが遺伝子のミューテーション[mutation](突然変異)によるとき、その遺伝子をホメオティック遺伝子[homeotic gene]、ホメオシスを示す個体をホメオティック変異体[homeotic mutant]という。
花の形成においては、ふつうの茎頂では葉に分化する原基(葉原基)が、外側から順に萼片・花弁・雄しべ・雌しべへと分化する。
1980年代末に、シロイヌナズナ(アブラナ科)のホメオティック変異体を使った研究に基づいて、花の形成過程に関する仮説が示された(ABCモデル)。
 シロイヌナズナ(アブラナ科)。春に道ばたで群生が時たま見られる小さな一年草だが、「モデル生物」[model organism]として多数の研究室で実験材料に使われ、遺伝子を通じた植物の理解の進歩に大きく貢献してきた。
シロイヌナズナ(アブラナ科)。春に道ばたで群生が時たま見られる小さな一年草だが、「モデル生物」[model organism]として多数の研究室で実験材料に使われ、遺伝子を通じた植物の理解の進歩に大きく貢献してきた。

シロイヌナズナの花は非常に小さいが、他のアブラナ科と同じ花の構成をしている。
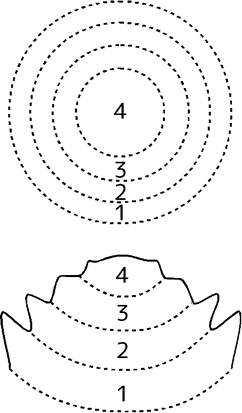
変異体でないシロイヌナズナ(野生型)の花の形成は、下の表のように説明できる。
| 領域 | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| はたらく遺伝子 | B | |||
| A | C | |||
| 葉原基→ | 萼片 | 花弁 | 雄しべ | 心皮 |
また、A~Cのいずれかがはたらかない変異体の花の形成でも、モデルどおりの結果となる。
| 領域 | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| はたらく遺伝子 | B | |||
| C | ||||
| 葉原基→ | 心皮 | 雄しべ | 雄しべ | 心皮 |
| 領域 | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| はたらく遺伝子 | ||||
| A | C | |||
| 葉原基→ | 萼片 | 萼片 | 心皮 | 心皮 |
| 領域 | 1 | 2 | 3 | 4・・・ | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| はたらく遺伝子 | B | B | B | ・・・ | |||||||
| A | |||||||||||
| 葉原基→ | 萼片 | 花弁 | 花弁 | 萼片・花弁の反復 | |||||||
ABCモデルは、単純なモデルでありながら、萼片・花弁・雄しべ・雌しべという順序で同心円状に並ぶことや、萼片・花弁・雄しべ・雌しべの間に連続性が見られる例、さまざまな奇形の成り立ちを説明する。
ABCモデルやABCモデルを改変したモデルは、シロイヌナズナ以外の多くの植物にも当てはまることが確かめられている。
| 領域 | 1 | 2 | 3 | 4 | |
|---|---|---|---|---|---|
| はたらく遺伝子 | B | ||||
| A | C | ||||
| D | |||||
| E | |||||
| 葉原基→ | 萼片 | 花弁 | 雄しべ | 心皮 | |
| 胚珠 | |||||
シロイヌナズナなどでは、さらに2つのクラスを追加した「ABCDEモデル」が立てられている。