5-1. 木本と草本
植物の生育方式の区分で日常的に使われるのは「木」と「草」だ。
木(木本 もくほん)
草(草本 そうほん)
木本のハリエンジュと草本のスズメノエンドウ、ともにマメ科
多くの植物は木本・草本のどれかに入るが、どのグループにも当てはまらないものもあり、2つのグループの性質を併せ持つものもある。寿命は長いが二次成長しない地上茎をもつタケ類(イネ科)やヤシ類(ヤシ科)は前者、一部の多年生つる性植物は後者だ。
5-1-1. 全形や生活史による区別: 木本
高木・低木・つる性木本
上左―高木のカツラ(カツラ科)
木本は、高さ(実際の高さではなく、潜在的に到達し得る高さ)とシュートの寿命によって高木と低木とに区別されるが、はっきりとした境界があるわけではない。やや低めの高木を「亜高木」、低めの低木を「小低木」「矮性低木」のように細分することもあるが、区別はさらにあいまいだ。また、他物に取り付いて伸びるつる性植物は、高さもシュートの寿命も(巻きつく対象など)条件次第で変わるので、高木と低木の区別どころか木本と草本の区別さえも意味をなさないことが多いので、ここでは別枠にする。
高木 低木 つる性木本(藤本)
植物体 最大樹高 シュート寿命 高木 自立 数m超 永続的 低木 数m以下 有限 つる性木本 他物に依存 可変 可変
常緑樹と落葉樹
高木・低木・つる性木本とも、一年のどの時期でも生葉がついている常緑樹 落葉樹
葉がつく枝
厚み
上面
葉脈
常緑樹
当年枝・前年枝等
厚い
暗緑色・光沢が強い
より不明瞭
落葉樹
当年枝のみ
薄い
明緑色・光沢が弱い
より明瞭
上: イヌシデ(カバノキ科)。落葉樹では当年枝だけに葉がある。 常緑樹タブノキ(クスノキ科─左)と落葉樹コナラ(ブナ科─右)の葉
常緑樹の葉の方が厚みがあって緑も濃く、葉脈が目立たず、上面の光沢が強い傾向があるが、区別し難いケースもある。
コナラ(左―ブナ科)の葉は紅葉または黄葉を経て、ヤマコウバシ(右―クスノキ科)の葉は黄葉を経て淡褐色の枯葉(褐葉)となり、枝に残る
枯葉があまり落葉せず、翌春まで残る種類もある。もちろん、落葉樹と見なされる。
落葉樹の葉の寿命は、定義からいって1年に満たない。常緑樹の葉は基本的に1年より長い寿命をもつが、種類によって大きく違う。クスノキ・ユズリハのように新葉が広がってからすぐに古い葉がいっせいに落ちる種類では、葉の寿命は1年をわずかに上回るに過ぎない。しかし、多くの常緑樹では葉はもっと長持ちで、2年枝や3年枝にも葉がついている。
左―クスノキ(クスノキ科)。春先に、新しい葉が開き始め、古い葉は紅葉・落葉する。 ヤブツバキ(ツバキ科)では、多くの葉が展開後3年目の初夏まで存続する。
常緑樹と落葉樹は常にくっきりと分かれるのではなく、さまざまな中間型があって、「半落葉性/半常緑性」と呼ばれることもある。低木やつる性木本の中には、気候によって常緑だったり落葉だったりするものがあり、冬を越す葉の有無は地域や年によって変わり、場合によっては個体や枝によって違うこともある。
キイチゴ類の低木ビロードイチゴ(バラ科)。3月、前年の葉の葉腋から、新しいシュート(当年枝)が伸び始める。同じ場所で、前年の葉が残っている枝(左)とほとんど枯れ落ちた枝(下)の両方が見られた。
ヤマツツジ(ツツジ科)は大形で楕円形の葉と小形で細長い葉の2タイプの葉をつける。大形の葉は秋に落葉し、小型の葉のみが越冬する。
ヤマツツジ(ツツジ科)
針葉樹と広葉樹
上―常緑針葉樹のヒマラヤスギ(マツ科)
裸子植物を針葉樹、被子植物の木本を広葉樹とよぶこともある。この名称は実際の葉の形状とぴったり一致するわけではなく、マツのように針状の葉をもつ裸子植物は多いが、鱗状の葉も少なくなく(ヒノキなど)、イチョウやナギ(イヌマキ科)のように幅広な葉もある。被子植物の木本でもサボテン類やガンコウラン(ツツジ科)などは針状の葉をもつ。
5-1-2. 全形や生活史による区別: 草本
生活環が一周する期間や発芽から開花・結実・枯死までの期間によって、一年草・二年草・多年草に分けられる。
一年草 二年草 多年草 一回繁殖型 多回繁殖型 発芽~結実・枯死 ~数ヶ月 1年超・2年未満 不定 発芽~次世代発芽 ~1年 2年 不定 開花・結実 1回(開花・結実後に枯死) 複数回
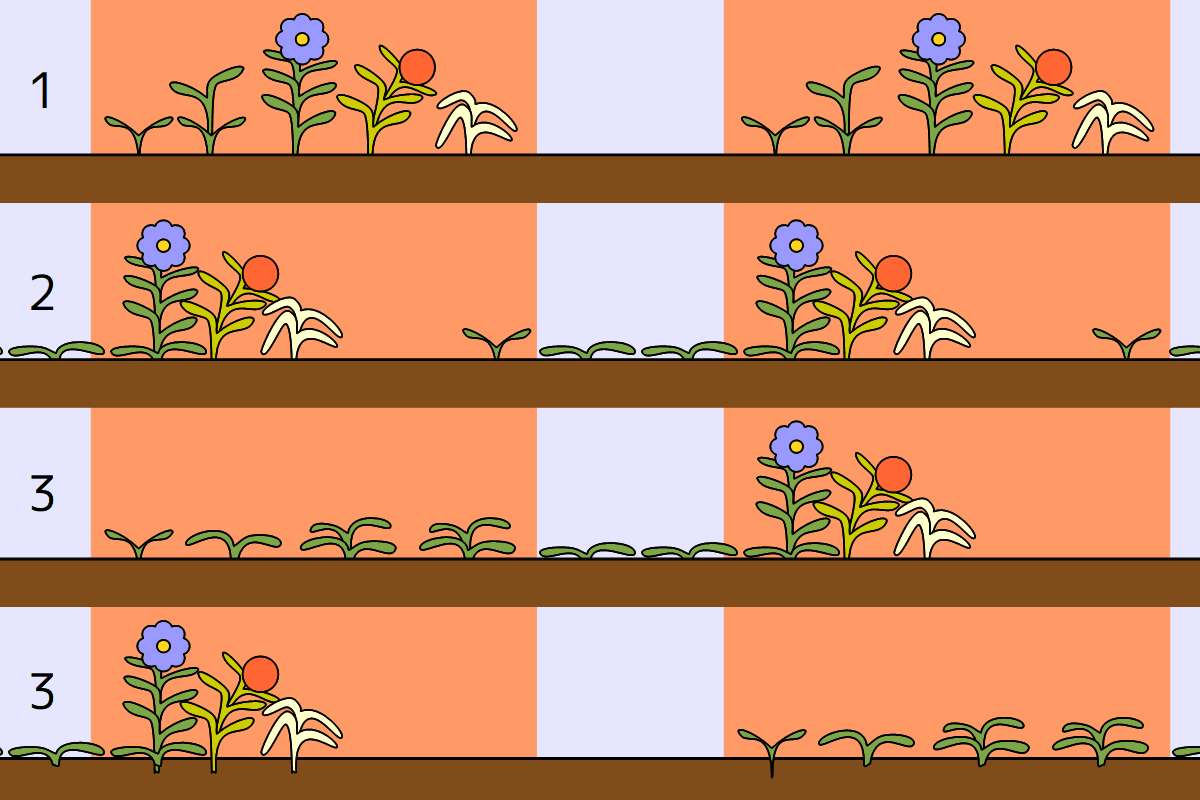
夏型一年草(1)・冬型一年草(2)・二年草(3)の生活環の模式図
一年草
夏型一年草[summer annual herb]: 春~夏に発芽して秋~冬までに枯死
冬型一年草または越年草[winter annual herb]: 秋~早春に発芽して初夏~夏までに枯死
周年型一年草: 発芽~枯死にはっきりした季節性がない
二年草 二年草のムラサキケマン(ケシ科)。晩春に散布された種子は、翌春に芽生え(上左)、光合成をして栄養を地下部に貯える(下左)。ロゼット葉を広げた状態で冬を越し(上右)、早春が来ると、越冬葉の上に新しい葉を広げ、花茎が伸びて開花(下右)、結実して種子を散布し、晩春には枯死する。
望ましくないが、冬型一年草(越年草)が「二年草」を呼ばれることもあり、紛らわしくないように二年草を「真正二年草」と呼ぶことがある。
多年草
一年草・二年草・多年草のどれにも、つる性のものがあり、「つる性一年草」「つる性二年草」「つる性多年草」のように表わされる。
多年草の生活史の多様性
多年草の多くは多回繁殖型[polycarpic]で、開花・結実しても枯死せず、条件さえ良ければ永続的に生き続ける。しかし、開花・結実をすると枯死する一回繁殖型[monocarpic]の種類も、ハマナデシコ(ナデシコ科)・マツヨイグサ類(アカバナ科)・オトコエシ(オミナエシ)・ボタンボウフウ(セリ科)など、少なくない。
海岸の斜面に生育する一回繁殖型多年草ボタンボウフウ(セリ科)は開花(左)・結実後に枯死する(右) 一回繁殖型多年草オトコエシ(オミナエシ科)の結実した個体は地下部まで枯死している
葉や茎の寿命を基準にすると、多年草は、常緑性・休眠性・半休眠性に区別できる。ただし、木本と同じでくっきりと区別できない場合もある。
常緑性: 一年のいつでも葉を付けている。常緑樹と同じく、常緑の草本でも葉の入れ替わり方はさまざまである。根元近くの茎が太く硬くなり、長い寿命を持って、木本の茎と同じような性質を持つ(木化)ものもある
休眠性: 冬・乾期などの生育不適期には、地上に出ているシュート系が枯れて地下部だけで過ごし、好適な条件になると再びシュート系を地上に広げる
半休眠性: 生育不適期には、地上のシュート系は地表に広がる葉を残して枯れて、ロゼット(地表の葉)と地下部で過ごす
ヤブラン(キジカクシ科)。春になると古い葉が倒れ伏している上から新しい葉が伸びてくる。 ノアザミ(キク科)の越冬するロゼット
木本と多年草のどちらとも言い切れない植物種は、少なくない。
木本と多年草の両面を備えている植物: コウヤボウキ(キク科)の茎は細くてあまり高くもならないが、2年間の寿命を持つ。ヤシ類(ヤシ科)やタケ類(イネ科)の茎は、太らないが寿命が長く、背も高くなる。
条件によって木本となる多年草: つる性の多年草に含められる植物では、条件によって茎の一部が木化して長い寿命を持つものがあり、ときには直径数センチまで太くなる(ボタンヅル属・ヘクソカズラ・クズなど→ツル植物の樹幹 )。
5-1-3. 木本の群集(1): 森林と低木やぶ
森林
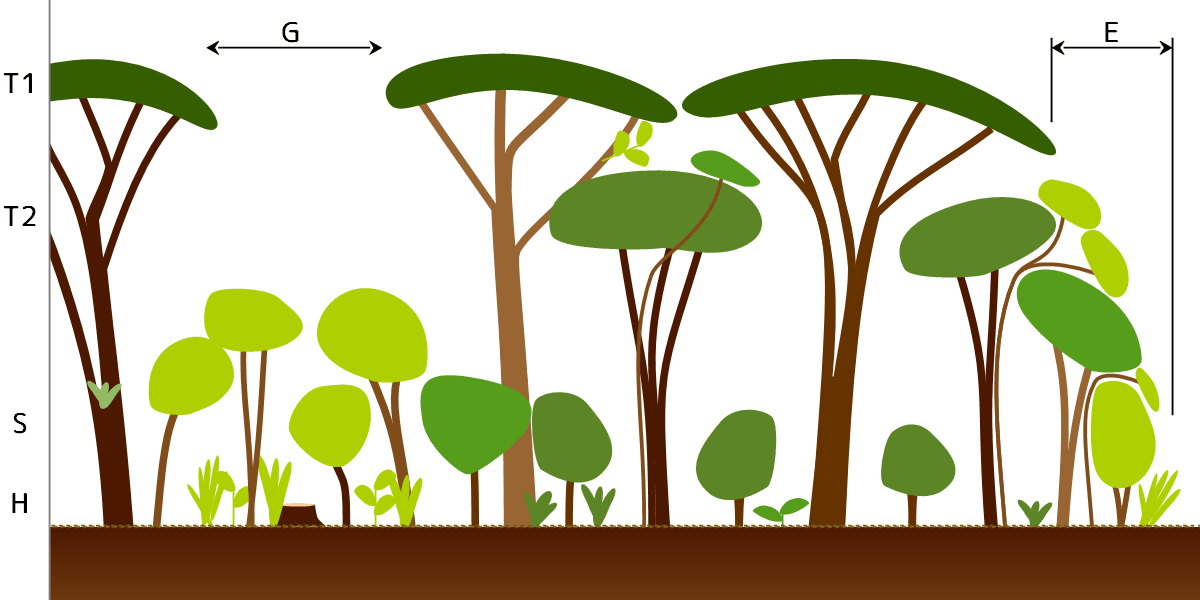
森林の断面(模式図)。右の矢印で示す範囲(E)が林縁、左寄りの矢印で示す範囲(G)が林冠ギャップ。
森林の中で、枝や葉は上から下まで均一に分布しているのではなく、枝や葉が集中する階層が見られる(階層化
下から見上げたシイ二次林。樹冠が石垣のようにぎっしりと並ぶ。
最上層で直接に太陽光を受ける枝・葉の集まりを林冠 [canopy; forest canopy]といい、階層としては林冠層[canopy layer]あるいは高木層[tree layer]という。林冠では枝葉の密度が最も高く、個々の木の枝葉の広がり(樹冠[tree crown])が敷き詰めたようにびっしりと並ぶ。
熱帯林では、林冠よりも上に樹冠が抜きんでている超出木/突出木[emergent tree]が点在することがあり、超高木層[emergent layer]を構成する。
横から見たシイ二次林 伐採により断面が露出したシイ二次林(福岡教育大学構内)。隣接地で伐採・造成されるシイ二次林の残土埋め立てのための伐採。 造成目的のシイ二次林の皆伐(福岡教育大学構内)。列状に刈り残された数本の木があって、普段は見ることができない森林構成木の樹形が見える。 左の2本がシイ・右の1本はコナラ。シイの方が林冠の葉は上下に狭い範囲に密集している。 林縁の木は、樹幹も樹冠も傾いている
林冠に届かない樹木は(下層木
シイ林の低木層では、シイの若い個体が上から洩れる弱い光を使って生育する シイ林の低木層で木洩れ日を受けるヒサカキ(ツバキ科|モッコク科)
下層木の一番下には、数十センチから人間の背丈を超える程度の低木層[understory; forest understory](図ではS)がある。
低木層の下、林内の地表(林床 [forest floor])近くの草本や幼樹を草本層と呼ぶこともある(図のH)。草本層は耐陰性のある草本(主に常緑の多年草)や木本からなるが、落葉樹林の場合は、晩秋~早春の落葉期に光合成をおこなう植物が見られることがある。
森林を構成する各層の出現種には違いがあるのがふつうだ。林冠の樹種は幼木あるいは若木として下層にも出現するが、アオキ・ヒサカキなど低木層以下しか出ない樹種もある。また、日本列島の冷温帯林は、低木層をササ類(イネ科)が高密度で占有する場所が多いのが大きな特徴だ。
林縁と林冠ギャップ
林のへり=森林が草地や空き地と接するところを林縁 [forest edge]といい、明るいところを好む樹種やツル植物が多く、林冠と同じように枝葉の密度が高くなっている。
カシ林の林縁。落葉樹とツル植物が目立つ。
林冠木が枯れたり倒れたりすると、その木の樹冠の分だけ林冠にすきま(ギャップ [gap; forest gap]または林冠ギャップ)ができる。森林の中に明るい空間が生じて、林縁と同じく明るいところを好む樹種や草本が茂る。樹木の生長や周囲の林冠の拡大によって、ギャップはゆっくりと小さくなり、消失にいたる。森林では、大小のギャップの出現と消失が繰り返されている。
シイ・カシ林で大きな倒木のところに行くと、上方にはさまざまな形・大きさのギャップが開いて、林床に光が差し込んでいる。 カラスザンショウ・ヌルデ・アカメガシワなどが生えている。いずれも、伐採跡や林縁など明るいところに多い樹種だ。 古い朽ち果てた倒木の上では、林冠ギャップはほとんど消えかけている。
低木やぶ(低木林)
森林が伐採された後の明るい場所に低木が密生するやぶ(低木やぶ)ができることがある。低木やぶは時間をかけて森林に移行していくが、土壌が不安定だったり(川べり、がけなど)、ひんぱんに刈り取りが行われるようなところ(道路ぎわなど)などでは、森林が発達しにくく、低木やぶが長期間保たれる場合もある。
伐採跡の低木やぶ
林床の低木と低木やぶの低木
同じ低木であっても、林床の低木層と低木やぶに共通する種類は少ない。林床の低木は伸長は遅いが寿命の長い茎を持つものが多く、対照的に、低木やぶの低木は伸長が速いが寿命が短い茎を次々と伸ばしていく傾向がある。
成長が速い樹木の茎は成長速度を稼ぐために丈夫さを犠牲にしており、材の密度が低く中心部の髄が大きくて切りやすい。
林縁や伐採地で速く成長する4樹種の枝(直径約1cm)の断面。順に、ネムノキ(マメ科)・ゴンズイ(ミツバウツギ科)・アカメガシワ(トウダイグサ科)・ニワトコ(スイカズラ科|レンプクソウ科)。
逆に、林床の低木や高木の幼樹は、丈夫だけれどゆっくりと成長する茎を持ち、材は緻密で髄も小さい。
林内でゆっくりと成長するヒサカキ(ツバキ科|モッコク科)と高木になるスダジイ(ブナ科)の枝(直径約1cm)の断面。
樹木の茎においては、成長速度と寿命(あるいは丈夫さ)の間に負の相関関係 正の相関関係
同じようなことは、葉でも成り立つ。常緑樹の葉は落葉樹に比べてクチクラ層が厚く、厚壁組織の量が多い傾向がある。反面、光合成に直接関与しない組織が多い分、光合成の効率は落ちる。
5-1-4. 樹木のシュートに見られるさまざまな戦略
最大成長速度: 最適な条件下でのどれくらい速く成長できるか
最大樹高: どれくらいの大木になり得るか=最適な条件下でどれくらいまで樹高を稼げるか
耐陰性: どれくらい弱い光の下で生存・成長を続けることができるか
これらは、どれも光を得る競争において優位となる特性だ。競争のスタートダッシュでは成長速度が、長期間の競争では最大樹高が、他の樹木の陰になった場合には耐陰性が生育を左右する。一方、三者の間には、負の相関関係がある。
最大樹高と耐陰性: 大木となるには、幹(主軸)が上方へと伸びていく方が有利だが、林床の木洩れ日で生育するのには、枝葉を横に広げる方が有利だ。限られた資源を前者では高く伸びること、後者では横に広がることに重点的に割り当てている。
最大樹高と成長速度: 丈夫で寿命の長い茎でないと大木になれない。しかし、限られた資源のもとでは、材を緻密にすることと成長速度は両立しない。
成長速度と耐陰性: 成長速度を稼ぐためには、光合成の効率の高い葉が有利だが、そのような葉は寿命が短く、暗いところでは作るのに要した資源を取り返す前に寿命が尽きてしまう。
このように、成長速度・最大樹高・耐陰性の3つのあいだには、
ある点で優位に立つと宿命的に別の点では不利になってしまう
という関係がある。このような関係を「トレードオフ
生物のさまざまな性質の間には、主に次の2つによる、無数のトレードオフの関係がある。
資源の制限[resource limitation]: 資源をある特性を高めることに割り当てると、他の特性のために使うことができる資源は当然のこととしてその分減ってしまう。成長速度・最大樹高・耐陰性の関係も資源の制限による。他に、次のような例が挙げられる。
生殖に多くの資源を割り当てる(例えば、多数の花、多数の果実をつける)と、生存や成長に割り当てられる資源の量が少なくなる。一年草は、生殖に全ての資源を割り当て、生存や成長を犠牲とする戦略をとる。ハマナデシコ(ナデシコ科)では、多くの個体は多数の花・果実をつけた後で枯死するが、花つきや結実が少なかった個体は越冬して翌年にも開花・結実する。 結実期のシマトネリコ(モクセイ科)。結実したシュートは、結実に多くの資源を費やし、葉の緑が早く褪せる傾向がある。
相互干渉[interference]: ある特性を持つことが、直接に他の特性を妨げる。次のような例が挙げられる。
目立つ花や実をつけて動物に花粉を運ばせたり(送粉)、果実を運ばせたり(散布)する植物の場合、葉や茎を茂らせて光合成量を高めると、花や実が葉の陰に隠れて受粉や散布の効率が落ちる(そのため、多くの植物では、花や実がつく枝では葉は退化して「花序」と呼ばれる「花専門の枝」になっている)
より目立つ花をつけて受粉・散布の効率を上げると、その分、花を食べる動物に発見される危険も増える。
多数の花が一斉に咲く方が、訪花昆虫に対して目立つ。しかし、同じ株の花粉が柱頭につく(自家送粉)の危険性が高まる
樹木のシュート系では、基本的な戦術(生態学では、「生活史戦略
戦略 シュートの特徴 有利な状況 伸長速度 寿命 伸長方向 開花サイズ 成長速度優先 早い 短命(1) 上方へ 小さい(2) 低木やぶ 樹高優先 遅い 長命 大きい(3) 森林の林冠 耐陰性優先 横に広がる 小さい(2) 森林の低木層 (1)新しいシュートを次々と出して寿命の短さをカバーする
上の3つを三角形の頂点とすると、頂点の中間に位置する戦略(中間戦略)や複数の戦略の使い分け(条件戦略)を考えることもできる。林冠を作るような高木は、耐陰性と樹高の両方を優先する中間戦略や、樹高が低く暗い光の下では耐陰性を優先し、樹高が高くなって直射光を受けるようになると樹高優先のシュート系を作るような混合戦略を採っていることが多い。
スダジイ(ブナ科)のシュートは、暗いところでは横方向に平面的に広がり、明るいところでは上方向へと伸びる。
また、つる性木本のように、他の植物や岸壁を身体を支えるのに使うことで、トレード・オフを「すり抜ける」植物もある。
成長速度・最大樹高・耐陰性優先のうち、どの戦略が有利は、環境によって大きく変わる。
森林が成立する過程(植生遷移
トレード・オフのため、どんなときでも優位に立つような(オールマイティーな)生物はない。このことによって生物群集は、さまざまな生態的地位(生態的ニッチ/ニッチ)[ecological niche]を占める個体群を含むようになり、地球レベルでは生物多様性[Biological diversity]を支えている。
ゲームやスポーツ、レースでも、トレード・オフ的な要素が入っていると戦術に多数の選択肢ができて、「深み」のある戦いになる。
先制型 長期戦型 特殊状況型 樹木 成長速度優先 樹高優先 耐陰性優先
バトル 素早さ+攻撃優先 攻撃+守備優先 素早さ+守備優先
レース 加速優先 最大速度優先 悪路走行性優先
攪乱とストレス
植物がぱらぱらと(まばらに)しか生えていない場所では、何らかの要因で植物の密度が抑えられている。Grime(1977)は、植物の繁茂を妨げる要因を攪乱[disturbance]とストレス[stress]に大別した。
要因 攪乱 ストレス 植物体や腐植が生態系から大量に除去される現象 成長を妨げる環境条件 特性 突発的 継続的 例 自然 人為 自然 人為 洪水・暴風・山火事 造成 日照不足/過剰 人工物による被陰
一言で言うと、攪乱は「突発的な破壊」、ストレスは「継続的な成長阻害」となる。
攪乱が全ての種にほぼ無差別に影響するのに対し、ストレスの影響はそれぞれの種が備える耐性によって異なる傾向が強い。
この点に着目すると、食害・病害や踏圧は攪乱ではなくストレスに分類される場合もあるのではないかと思う。
CSR三角形
攪 強 R 中 弱 C S 弱 中 強 ストレス
右の図は、ストレス・攪乱の強さと植被率の関係を表わしたもので、色の濃いところほど高い植被率を示す。ストレスと攪乱の両方が強いところは、植生じたいが成立しないので、空白となる。
Grimeは、三角形の3つの頂点に生育する植物の特性に注目することで、植物の生活史戦略の多様性を説明しようと試みた。
C―競争力植物: 光・水・養分の量に応じてシュートと根の比率を変える。水・養分の吸収が速く、好条件下で速い成長。
R―攪乱依存植物: 早めに開花結実し、貯蔵用分の多くを種子に割り当てる。種子繁殖後に枯死することが多い。
S―ストレス耐性植物: 条件にかかわらずゆっくりと成長。
上の話に当てはめると、成長速度優先はR、樹高優先はC、耐陰性優先はSになる。
Grime JP (1977) Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory.
American Naturalist 111(982):1169–1194. doi:10.1086/283244
5-1-5. 木本の群集(2): 常緑樹と落葉樹
日本列島では、落葉樹が晩秋から早春にかけて休眠状態に入って越冬する。これに対して、休眠できない多くの常緑樹は温暖な地域に分布が限られてしまう。しかし、温暖な地域、例えば南西諸島にも落葉樹は分布しているし、東北や北陸の積雪地帯に分布する常緑樹もある。だから、寒さだけでは両者の分布を説明することができない。
常緑林ができるような温暖な気候下で、落葉樹が生育するのは、次のようなところである。
低木やぶ: 低木やぶでは、気候にかかわらず、ほとんどの種類が落葉性である。
林の縁のような明るいところ
沢筋のように土壌が不安定なところ(極端に不安定な場合は、森林ではなく低木やぶになる)
城山の一部を自然教棟の西端から撮影したもの。上が紅葉(12/01)、下が新緑(4/26)の季節。中央の沢に沿ったところや、右下の林の縁に落葉樹が多い。 林床をツルミヤマシキミ・ヒメアオキ・ヒメモチ・エゾユズリハが占める日本海側多雪地のブナ林
積雪地帯では、落葉林の下層木として常緑の低木が見られる。これらの木は、冬の間積雪によって極端な冷え込みから守られている。常緑樹のアオキ・ミヤマシキミ・イヌガヤ・カヤなどは、雪が少ない地方では大型の低木や高木になるが、日本海側の多雪地帯では他の地方でよりも背が低く、冬の間は積雪に埋没する。
ヒメアオキ(ミズキ科|ガリア科)。雌雄異株の常緑低木。アオキの変種で、日本海側の多雪地帯に分布する。
5-1-6. 草は木よりも多様な場所に生えている
草の生育する範囲は木よりも広い。森林で高木と共存するものもあれば、低木やぶに混じるものもある。寒冷・積雪・不安定な土壌・養分が少ない土壌・過剰な土壌水分・人間による刈り取り・火入れなどは、限度を超えると森林はもちろん低木やぶの成立も妨げる。しかし、このようなところでも裸地であることは少なく、大抵は草が茂っている。水生植物のように、特殊な生育地に生育しているものもある。
常緑の多年草は落葉樹林や常緑樹林の林床 ホソバナコバイモ(ユリ科)。左は4月はじめの開花期、右は5月中旬の結実期ですでに葉が茶色になっている。種子散布のあとは10ヶ月近い長い休眠に入る。
林のへり(林縁 ギャップ
自然条件や人間の手によって、頻繁に破壊されるような生育地では一年草が多くなる。
テキスト目次に戻る ホームに戻る

 木本のハリエンジュと草本のスズメノエンドウ、ともにマメ科
木本のハリエンジュと草本のスズメノエンドウ、ともにマメ科


 上左―高木のカツラ(カツラ科)
上左―高木のカツラ(カツラ科)









 キイチゴ類の低木ビロードイチゴ(バラ科)。3月、前年の葉の葉腋から、新しいシュート(当年枝)が伸び始める。同じ場所で、前年の葉が残っている枝(左)とほとんど枯れ落ちた枝(下)の両方が見られた。
キイチゴ類の低木ビロードイチゴ(バラ科)。3月、前年の葉の葉腋から、新しいシュート(当年枝)が伸び始める。同じ場所で、前年の葉が残っている枝(左)とほとんど枯れ落ちた枝(下)の両方が見られた。




 夏型一年草(1)・冬型一年草(2)・二年草(3)の生活環の模式図
夏型一年草(1)・冬型一年草(2)・二年草(3)の生活環の模式図












 横から見たシイ二次林
横から見たシイ二次林 伐採により断面が露出したシイ二次林(福岡教育大学構内)。隣接地で伐採・造成されるシイ二次林の残土埋め立てのための伐採。
伐採により断面が露出したシイ二次林(福岡教育大学構内)。隣接地で伐採・造成されるシイ二次林の残土埋め立てのための伐採。

 林縁の木は、樹幹も樹冠も傾いている
林縁の木は、樹幹も樹冠も傾いている
 シイ林の低木層で木洩れ日を受けるヒサカキ(ツバキ科|モッコク科)
シイ林の低木層で木洩れ日を受けるヒサカキ(ツバキ科|モッコク科)





















 林床をツルミヤマシキミ・ヒメアオキ・ヒメモチ・エゾユズリハが占める日本海側多雪地のブナ林
林床をツルミヤマシキミ・ヒメアオキ・ヒメモチ・エゾユズリハが占める日本海側多雪地のブナ林
