 カキノキでは、果肉の一部がぬるぬるしたゼリー状になって種子を完全に包む(簡単には取れない)。カキノキのような比較的大形の種子では、鳥が種子を除けて果肉だけを食べるリスクがあるので、この特徴は、鳥が種子を丸呑みすることを促している可能性がある。
カキノキでは、果肉の一部がぬるぬるしたゼリー状になって種子を完全に包む(簡単には取れない)。カキノキのような比較的大形の種子では、鳥が種子を除けて果肉だけを食べるリスクがあるので、この特徴は、鳥が種子を丸呑みすることを促している可能性がある。鳥や哺乳類の糞は、しばしば植物の果実・種子を含んでいる。


 大学内の路上やコンクリートの上では、(おそらく)テンの糞をよく見かける。5月終わり頃はヤマザクラとクサイチゴの果実がたくさん入っている。
大学内の路上やコンクリートの上では、(おそらく)テンの糞をよく見かける。5月終わり頃はヤマザクラとクサイチゴの果実がたくさん入っている。

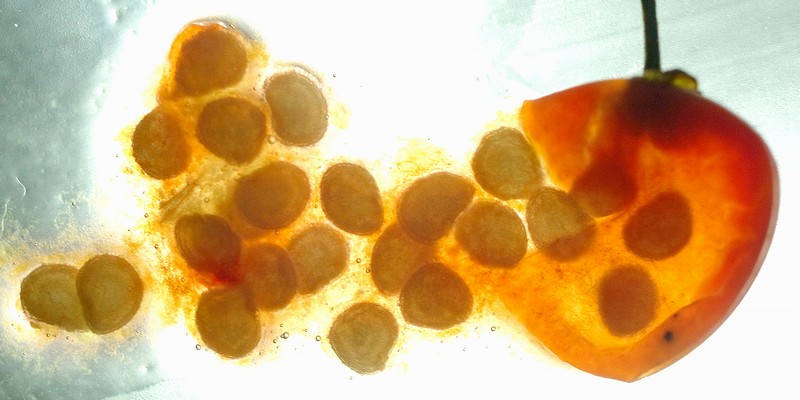
 上左―山頂の展望台にあった鳥の糞(おそらく複数回分)には、さまざまな植物の種子が入っていた
上左―山頂の展望台にあった鳥の糞(おそらく複数回分)には、さまざまな植物の種子が入っていた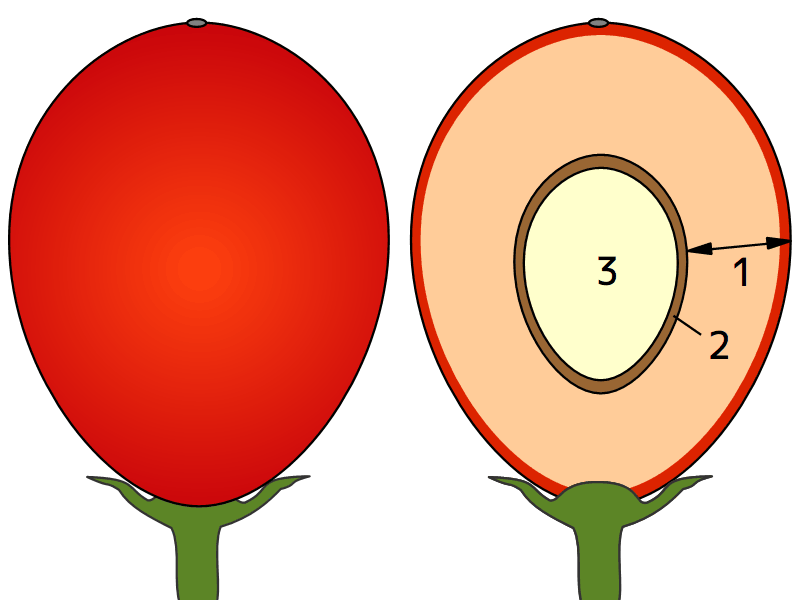
鳥獣に食べられてから糞として排泄されることで運ばれる果実・種子の多くは、次の3つの部分からできている(外側→中心の順)。
「可食部と保護部が花のどの部分から発達するか」には色々あって、糞散布体の多様な起源を示すとともに、糞散布体の重要な分類基準となっている。
| 果実の名称 | 花被 | 花托 | 花托筒 萼筒 |
子房壁 | 種衣 | 種皮 | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 漿果 | × | 可食部 | なし | 保護部 | 上位子房由来 | |||||
| × | 可食部 | 下位子房由来 | ||||||||
| ナシ状果 | ||||||||||
| 核果(石果) | × | 可食部 | 保護部 | 薄皮として残存 | ||||||
| バラ状果 | × | 可食部 | 保護部 | 周位の花由来 | ||||||
| イチゴ状果 | × | 可食部 | × | 雌しべ複数 | ||||||
| (クワなど) | 可食部 | × | ||||||||
| ― | × | 裂開または 非裂開 |
可食部 | 保護部 | ||||||
| なし | 可食部 | 保護部 | ||||||||
糞散布の果実・種子の色は黄・赤・紫・黒紫・黒(ほとんどは濃い黒紫)などさまざまで、特に赤と黒紫~黒が多い。また、黒熟する果実には、サクラのように途中段階で赤を経るものが多く見られる。













糞散布に代表される動物被食散布は、動物送粉と並んで、被子植物と動物との相利関係だ。共進化の結果として、動物媒花と同様に、糞散布体は、視覚・嗅覚への信号によって動物を誘引し、動物に可食部という報酬を与える。動物によって散布距離・種子生存率・散布指向性に優劣がある場合、糞散布体の特徴によって優れた散布動物が選別される場合もある。また、糞散布体の特徴には、散布動物の行動を制御していると考えられるものもある。
移動距離が長く食物を丸呑みする鳥類は潜在的に優秀な散布者だ。鳥類は四色型色覚をもち、緑色の葉に混じる赤い果実を識別する能力が高い。一方、昆虫の多くは赤の感受性が低く、多くの哺乳類は二色性色覚で、赤と緑を識別しづらい。哺乳類の中で、サル類の一部は果実食に適応する過程で三色型色覚が進化し、緑と赤を見分けることができるようになった。

トウガラシ(ナス科)は、果実の辛味成分(カプサイシノイド; カプサイシンとカプサイシン類似物質の総称)が食害者排除と散布者選別にはたらいていることが推定される有名な例だ。
カプサイシノイドは抗菌作用を示し、カメムシが吸汁したトウガラシの果実に吸汁痕から侵入する菌類(Fusarium)の増殖や種子への感染を抑制する効果がある。カメムシの穿孔頻度が低い地域では、カプサイシノイドを殆ど含まないトウガラシが分布する。一方、カプサイシノイドは哺乳類の高熱や酸を感知する受容体タンパクTRPV1(VR1)を刺激して辛味感覚を引き起こすが、鳥類のTRPV1はカプサイシノイドに対する感受性がないため、辛いトウガラシは哺乳類に好まれず、鳥類に独占されることになる。
トウガラシのように、可食部の二次代謝産物に散布者を選別するはたらきがあるという仮説を、指向性抑止仮説[Directed deterrence hypothesis; DDH]という。
カキノキでは、果肉の一部がぬるぬるしたゼリー状になって種子を完全に包む(簡単には取れない)。カキノキのような比較的大形の種子では、鳥が種子を除けて果肉だけを食べるリスクがあるので、この特徴は、鳥が種子を丸呑みすることを促している可能性がある。

人間が花を観賞し、蜂蜜を利用するのは、動物媒花と送粉動物の相利関係に便乗した結果と見なすことができる。同じように、糞散布植物と散布動物の相利関係に便乗するかたちで、人間は、糖分やビタミンに富んだ可食部と鮮やかな色を持つ糞散布体を、品種改良を通じて、「果物/フルーツ」として食用・観賞用に利用する。この点でもトウガラシは大変興味深い例で、カプサイシノイドの抗菌作用と刺激性の両面がともに活用されてきた(真紅の果皮も観賞用に利用されている)。

子房壁から果肉ができ、種皮が硬い殻の役目をするを漿果または液果という。
カキノキ(カキノキ科)、ブルーベリー・コケモモ(ツツジ科)、ブドウ(ブドウ科)・トマト・ナス・ピーマン(ナス科)・バナナ(バショウ科)・キウイフルーツ(マタタビ科)・スイカ・キュウリ(ウリ科)、ビワ・リンゴ・ナシ(バラ科)、アボカド(クスノキ科)など。



子房下位の果実では、子房は花托に埋め込まれているため、子房壁と花托の組織が融合して果皮になる。この場合も、ふつうは漿果と呼ぶ。




漿果・液果は可食部に包まれた裂開しない果実の総称として使われることがある。また、漿果・液果のうち、身近で目立つ特徴のあるものに個別の名称がついていることがある。
下位子房のビワ・リンゴ・ナシ(バラ科)では、果実の断面を見ると花托筒から発達した部分が果肉の多くを占めている。このため、特に「ナシ状果[pome]」と呼んで区別することもある。


 リンゴ果実の横断面と花・果実縦断面・果実横断面の模式図。横断面では、花柱につながる心皮背束(●)✕5・花弁につながる維管束(●)✕5・萼片につながる維管束(◎)✕5を示す。
リンゴ果実の横断面と花・果実縦断面・果実横断面の模式図。横断面では、花柱につながる心皮背束(●)✕5・花弁につながる維管束(●)✕5・萼片につながる維管束(◎)✕5を示す。
 ミカン属の果実横断面(部分)の模式図
ミカン属の果実横断面(部分)の模式図
柑橘類の漿果では、子房室を囲む内果皮(瓤嚢/じょう嚢; じょうのう)の内壁から多数の突起が成長して果汁を貯える袋(砂瓤/砂じょう; さじょう)となる。食用部が果皮のごく一部からできるので「ミカン状果」[hesperidium]と呼んで区別することが多い。


子房壁のうち、外側の部分は果肉となり、内側の部分が種子を包む硬い殻となる。種皮は薄い。種子+硬い殻を「核」[kernel]という。
ウメ・モモ・サクランボ(ともにバラ科)・オリーブ(モクセイ科)・マンゴー(ウルシ科)など。





サクランボ・ウメ・モモ・マンゴーの「タネ」から果肉を完全に取るのは面倒だ。これは、「タネ」=核の組織が果肉とつながっているためだ。本物の種子は核の中に入っていて、核をこじ開けたり砕いたりすればきれいに取れる(モモやウメでは「仁」とか「天神さま」とか呼ばれているもの)。アーモンドもモモ・ウメに近い種類の種子で、「仁」とよく似ている。表面の茶色で筋のある薄皮が種皮だ。


 モモ(バラ科)の核と2つに割ったところ。中にアーモンドとよく似た種子が1個入っている。
モモ(バラ科)の核と2つに割ったところ。中にアーモンドとよく似た種子が1個入っている。核の中に複数の種子が入っているものも、決して珍しくない。


バラ科の糞散布果実は形態の多様性が高い上に、多くの食用果実が含まれていることから、固有の名称がつけられているものがある。上で出てきた、液果(漿果)の一型と見なせるナシ状果もその一つだ。



子房周位の花に由来し、子房を取り囲む花托筒(萼筒)が果肉となる果実は、バラ属・ピラカンサなどバラ科の一部に見られ、「バラ状果」[cynarrhodium]と呼ばれる。果実を割ると、中の空間にまわりの組織から独立した雌しべ由来の部分がある。同じバラ科のナシ状果(ビワ・リンゴ・ナシ)は、「子房がまわりの組織から独立している/いない」を除けば、このタイプと似ているところもある。
ナガバモミジイチゴやクサイチゴ、ラズベリー・ブラックベリーなどのキイチゴ類(バラ科)のような、多数の雌しべが核果になった核果の集合果は「筋子」のような外見で、キイチゴ状果と呼ぶことがある。

 クサイチゴ(バラ科)の熟した果実。透き通った赤い粒が集まっている。粒の1つ1つは、受粉した雌しべが成長したもの。受粉の成功率によって、大きさが違ってくる。赤い粒を拡大すると、残存する花柱が見える。1つ1つがウメ・モモ・サクランボの果実(核果)に相当するもので、赤くて透明な果肉の中に核が透けて見える。受粉しなかった雌しべも回りにへばりついている。
クサイチゴ(バラ科)の熟した果実。透き通った赤い粒が集まっている。粒の1つ1つは、受粉した雌しべが成長したもの。受粉の成功率によって、大きさが違ってくる。赤い粒を拡大すると、残存する花柱が見える。1つ1つがウメ・モモ・サクランボの果実(核果)に相当するもので、赤くて透明な果肉の中に核が透けて見える。受粉しなかった雌しべも回りにへばりついている。 粒がついている部分(花托)は、すかすかですきまだらけ。
粒がついている部分(花托)は、すかすかですきまだらけ。イチゴ・ヘビイチゴ(バラ科)などで、キイチゴ類と同様に花は雌しべをたくさん持っている。果肉は花托がふくらんでみずみずしくなったもので、果肉の表面についている多数のゴマ粒のようなものが子房だ。


 ヤブヘビイチゴ(バラ科)の果実
ヤブヘビイチゴ(バラ科)の果実





受粉の後も花被が残って厚くなり可食部になる。クワ(クワ科)・イチジク(クワ科)・ツルムラサキ(ツルムラサキ科)・ドクウツギ(ドクウツギ科)など。







イチジクでは、つぼのような花序の内側に多数の花がついている。つまり、イチジクの実と呼ばれるのは一つの花序、中の食べる粒々の一つ一つが花で、つぼ状の部分の内側もやわらかい可食部となっている。日本で栽培しているイチジクは果実が熟さないタイプのものだが、中近東などから輸入されている「乾しイチジク」は、果実が熟しているせいでじゃりじゃりとした歯応えがある。
果実をこじ開けると、中に可食部をまとった種子が詰まっている。
熟すると果皮が裂開して種子(と可食部)が露出することが多いが、ムベ(アケビ科)のように熟しても開かないものもある。熱帯果実のレイシ・ランブータン(ともにムクロジ科)・パッションフルーツ(トケイソウ科)・マンゴスチン(テリハボク科)・ドリアン(アオイ科)も非常に堅い果皮におおわれたまま熟する(サルのような哺乳動物がこじ開けて食べるのではないかと言われている)。



 トベラの実。熟するとぱっくりと開いて、赤くてねばねばした納豆のような種子が露出する。
トベラの実。熟するとぱっくりと開いて、赤くてねばねばした納豆のような種子が露出する。




冬眠や冬前の「食いだめ」をせずに冬を越す小形哺乳類(ネズミ・リスなど)・小形~中形鳥類(カケスなど)は、秋の間に食物を地中や岩の隙間、樹幹の割れ目に貯え、冬季の食料源とする習性をもつ(貯食)。
ドングリ(コナラ属の果実の総称)・ブナ・シイ(ブナ科)やクルミ属(クルミ科)など秋に熟する比較的大型の堅果(種子1個が硬い殻に覆われた果実)は、貯蔵場所に運ばれ、食べ残されたり食べ忘れられて発芽することがある(貯食型散布)。

比較的小型の果実・種子の中には、可食部が外側を覆うのではなく、果実・種子の本体に可食部が付着しているという感じになっていて、アリの巣に運び込まれ、そこで可食部を切り離されて、本体は巣のまわりや巣中のゴミ捨て場に放棄されるものがある。このような散布はアリ散布と呼ばれる。シソ科の一部・スゲの一部などでは果実に、スミレ属・スズメノヤリ・キケマン属などでは種子にアリ散布用の可食部がついている。


 ムラサキケマン(ケシ科)。熟した果実は瞬間的に裂けて黒い種子を弾き飛ばす(右上)。種子には白い小さな附属体がついていて(右下)、地上に落ちてからアリによって持ち去られる。 |
  |



