7-1. 被子植物の有性生殖
7-1-1. 有性生殖生物の生活環

有性生殖生物の生活環[life cycle]では、次のように、受精と減数分裂を通じて単相(n)世代と複相(2n)世代が繰り返される(世代交代)。
(1) 受精[fertilization]
ゲノム1組("n"と略される)を持った雄性配偶子(♂配偶子)[male gamete]または精細胞[sperm cell]と、やはりnを持った雌性配偶子(♀配偶子)[female gamete]または卵細胞[egg cell]とが融合して、ゲノム2組("2n"と略される)を持った受精卵ができる。
-
種子植物(イチョウとソテツをのぞく)では、雄性配偶子は花粉粒に入ったまま卵の近くまで運ばれ(送粉)、花粉粒から伸びた突起(花粉管)によって卵細胞まで運ばれる(花粉管受精)。
-
イチョウとソテツでは、花粉粒が送粉によって運ばれ、花粉管も伸びる。その後、花粉管から精子が放出されて短い距離を泳いで卵細胞に達する(花粉管受精+精子受精)。
-
シダ植物・コケ植物では、雄性配偶子は小さくて鞭毛を持っていて水中を泳ぐことができ(遊泳性がある雄性配偶子は「精子」[sperm]という)、雌性配偶子は大きくて移動力がない。精子が水中を泳いで卵細胞に達して受精が起こる(精子受精)。
-
藻類には、両方の配偶子の両方が鞭毛を持っていて移動するものが多数を占める他、精子受精をするもの、両方の配偶子とも鞭毛を持たず、細胞が接したときに融合して受精が起こるものなど、さまざまなタイプがある。
(2) 複相世代(2n世代・胞子体世代)
受精卵が体細胞分裂を繰り返して2nをもった多細胞体となる。植物では胞子体[sporophyte]と呼ぶ。


タマネギ(ユリ科|ネギ科)の根端の中期染色体。タマネギの複相世代(胞子体)の細胞は16本の染色体を持つ(このことを、「2n=16」と表現する)。左は、くびれ(動原体)が中央にあるもの→端近くにあるもの、の順に並べたもので、「形が良く似た染色体の対×8」から構成されていることが推測できる。対になった染色体のうち、1つは母親から胚珠を通じて、もう1つは父親から花粉を通じて受け継いだものだ。
(3) 減数分裂[meiosis]
2nの多細胞体(植物では胞子体)の一部の細胞が減数分裂をする。減数分裂の初めに、対になった染色体の間でつなぎ換えが起こり、父親からの染色体と母親からの染色体が混じり合う。1個の2n細胞(植物では胞子母細胞)が減数分裂すると、4個のn細胞(胞子[spore])ができる。
(4) 単相世代(n世代・配偶体世代)
減数分裂で出来たnの細胞(植物では胞子)が体細胞分裂を繰り返してnを持った多細胞体となる。植物では配偶体[gametophyte]という。配偶体の一部の細胞が雄性配偶子と雌性配偶子になる。
このように受精と減数分裂を通じて単相(n)世代と複相(2n)世代を繰り返すこと(世代交代)によって遺伝的な組み替えが行われる。
「胞子」
胞子[spore]の元々の定義は「空中を飛散または水中で分散する耐久性の繁殖単位」で、植物・菌類・変形菌・細胞性粘菌・卵菌・動物(刺胞動物門ミクソゾア亜門)・バクテリアなど、さまざまな生物群でつくられ、有性生殖の過程でできるものも無性生殖でできるものもある。
バクテリアの胞子[bacterian spore]は「芽胞」あるいは内生胞子[endospore]と呼ぶことが多い。
上の定義が当てはまる胞子は、コケ植物とシダ植物では減数分裂によってつくられる。種子植物で減数分裂でできる細胞は、元々の定義を満たさないが、生活環の記述に統一性を与えるため、定義を拡張して「胞子」と呼ぶ。
7-1-2. 生活環の多様性: 雌雄の分化

右の図は、多くのシダ類に見られる生活環だ(下表のI型)。上半分が2n世代、下半分がn世代で、円/楕円は単細胞体、六角形は多細胞体を示す。シダ類では配偶体は「前葉体」[Prothallus]と呼ばれる。
植物に見られるさまざまな生活環のうち、このタイプは、(1)胞子体・(2)胞子母細胞と胞子・(3)配偶体のいずれも雌雄に分かれず(同型)、雌雄があるのは配偶子だけで、雌雄の分化という点では最も単純だ。
植物全体では、各段階で次のように雌雄が別々になる生活環がみられる。
-
配偶体が雌雄に分かれる(コケ植物では雌雄異株)
-
胞子母細胞・胞子・配偶体が雌雄に分かれる
- 雌―大胞子母細胞・大胞子・雌性配偶体
- 雄―小胞子母細胞・小胞子・雄性配偶体
-
胞子体・胞子母細胞・胞子・配偶体が雌雄に分かれる(雌雄異体・雌雄異株)
生活環と雌雄性の多様性
| 型 |
グループ |
胞子体 |
胞子母細胞・胞子 |
配偶体 |
配偶子 |
| 雌性 |
雄性 |
雌性 |
雄性 |
雌性 |
雄性 |
雌性 |
雄性 |
| I |
シダ植物の多く |
両性個体
|
胞子母細胞
胞子
|
両性前葉体 |
卵
細
胞 |
精
細
胞 |
| II |
コケ植物の多く |
両性胞子体
(雌個体上) |
雌個体 |
雄個体 |
| シダ植物の一部 |
両性個体
|
雌性
前葉体 |
雄性
前葉体 |
| III |
大胞子
母細胞
大胞子 |
小胞子
母細胞
小胞子 |
雌雄同株の
種子植物 |
胚嚢 |
花粉粒 |
| IV |
雌雄異株の
種子植物 |
雌個体 |
雄個体 |
植物の生活環は、雌雄分化の程度によって上の表のような4つの型に分けられる。冒頭で挙げた、多くのシダ植物の生活環(雌雄分化は配偶子のみ)は表のI型に当たる。

[II型] コケ植物の多くやシダ植物の一部(スギナなど)では、配偶体が雌雄に分かれており、胞子から雌性配偶体か雄性配偶体のどちらかができる(右図)。コケ植物の場合、生活の単位は配偶体なので、雌雄異体(雌雄異株)となる。

[III型] 種子植物とシダ植物の一部(クラマゴケ類・水生シダなど)では、胞子にも♀胞子(ふつう大形なので大胞子[megaspore])と♂胞子(ふつう小形なので小胞子[microspore]とがある(右図)。大胞子から雌性配偶体、小胞子から雄性配偶体ができる。種子植物では、雌性配偶体=胚嚢、雄性配偶体=花粉粒で、胞子体につく有性生殖器官に組み込まれている。

[IV型] 種子植物の中には、右図のように、胞子体が雄と雌に分かれるものもある(雌雄異体・雌雄異株)。♀胞子体(雌個体・雌株)は大胞子を作り、♂胞子体(雄個体・雄株)は小胞子を作る。
配偶体または胞子体が省略された生活環
陸上植物の生活環には配偶体(n)と胞子体(2n)の2種類の多細胞体があるが、他の多細胞生物では、一方がスキップされる生活環もある。

多細胞動物や藻類の一部(褐藻のヒバマタ目・車軸藻類など)では、減数分裂でできた細胞は(体細胞分裂をすることなく)そのまま配偶子(卵と精子)となる。多細胞体は受精卵から発生する個体(2n)のみで、多細胞動物では雌雄に分かれる雌雄異体が多く、藻類では雌雄同体がふつうだ。右図は雌雄異体の場合を示す。

多細胞動物のうち、一部の昆虫(ハチ目など)やワムシ類では、卵が受精すると雌個体(2n)となり、受精しないと雄個体(n)となる性決定様式が進化し、半数体(n)と二倍体(2n)の多細胞体の両方が存在するようになった。
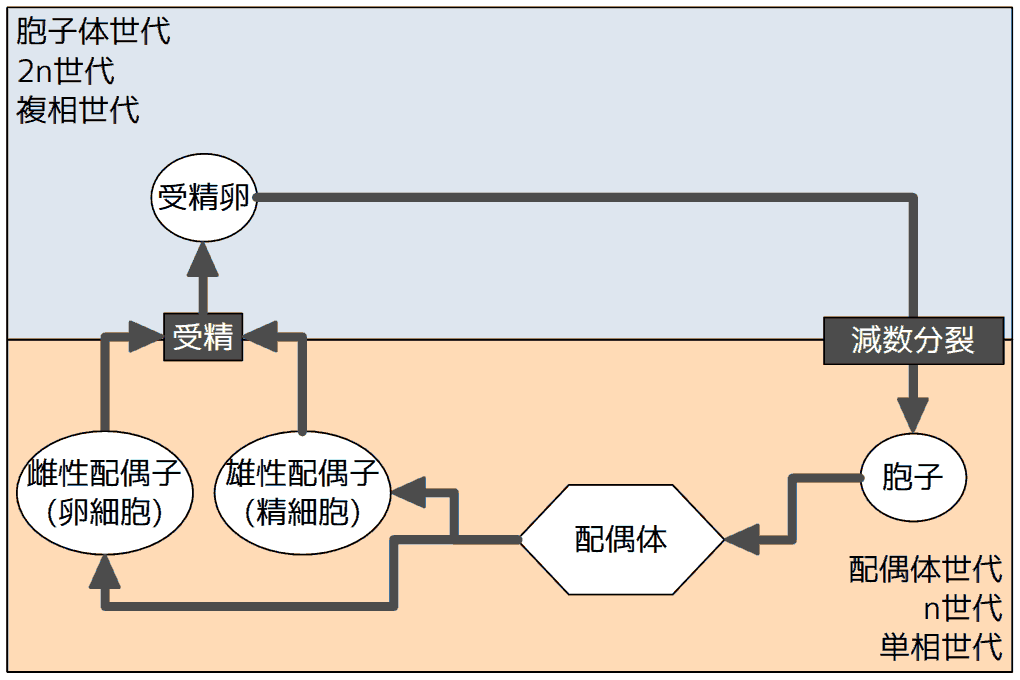
逆に、藻類には、胞子体がなく受精卵が(体細胞分裂をすることなく)減数分裂をして胞子をつくるもの(右図)もある。
7-1-3. 生活環の多様性: 配偶体と胞子体の関係
藻類では両方がほとんど同じ大きさ・形のものもある(アオサ類など)が、陸上植物では胞子体と配偶体の大きさ・形に大きな差がある。

 上─ギンゴケ(コケ植物・セン類)。びっしり並ぶ配偶体(茎葉体)の一部が先端に長い柄のついた胞子体をつける。
上─ギンゴケ(コケ植物・セン類)。びっしり並ぶ配偶体(茎葉体)の一部が先端に長い柄のついた胞子体をつける。
左―マキノゴケ(コケ植物・タイ類)の青黒く平たい配偶体(葉状体)と、そこに形成されているマッチ棒状の胞子体(マッチ棒の頭が胞子嚢)。
コケでは、茎や葉のように見える部分(茎葉体や葉状体)は配偶体で、胞子をつくるときだけ、胞子体が配偶体上に形成される。
シダの配偶体は「前葉体」[prothallus]と呼ばれる小さな植物体で、ハート形の葉状体が多いが、不定形や塊状のものもある。配偶体の裏面にできた卵細胞が受精して受精卵になり、大きな胞子体へと成長する。
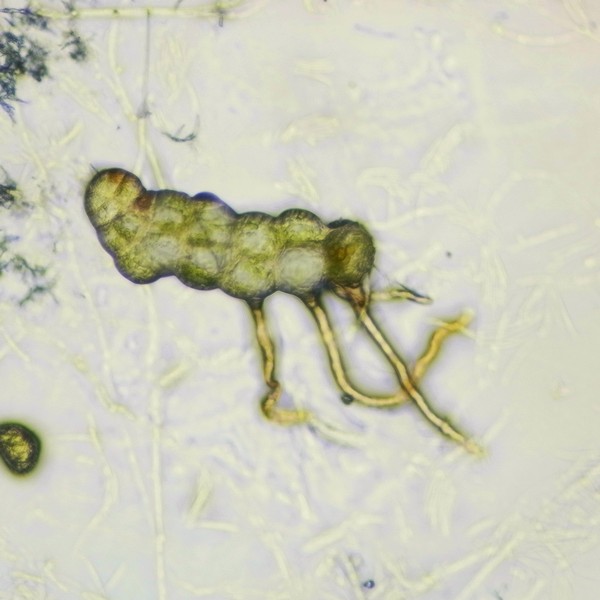
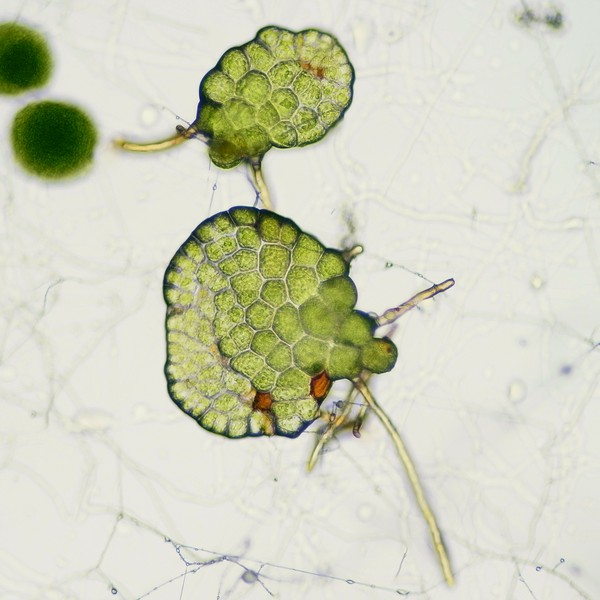
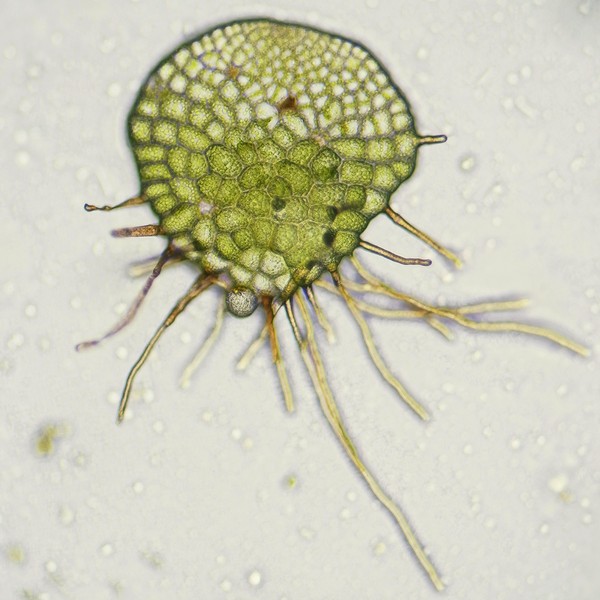
マメヅタ(ウラボシ科)の胞子から発生した若い前葉体


 ミゾシダの前葉体(上2枚)と胞子体(左)。前葉体はコケ植物の葉状体と同様、茎と葉の区別がなく、維管束や根をもたず、仮根で基質に付着する。細胞層は1層で、細胞間隙がないために透き通った緑色をしている(写真では、やや暗い緑色に見える)。上左の前葉体では、裏側から幼い胞子体が伸びだしており、質感の違いが分かりやすい。
ミゾシダの前葉体(上2枚)と胞子体(左)。前葉体はコケ植物の葉状体と同様、茎と葉の区別がなく、維管束や根をもたず、仮根で基質に付着する。細胞層は1層で、細胞間隙がないために透き通った緑色をしている(写真では、やや暗い緑色に見える)。上左の前葉体では、裏側から幼い胞子体が伸びだしており、質感の違いが分かりやすい。
 シダの前葉体は湿った土が露出しているような場所が見つけやすい。
シダの前葉体は湿った土が露出しているような場所が見つけやすい。
 種子植物の配偶体はさらに小さく、有性生殖器官(いわゆる「花」)の内部に作られる。♀配偶体は胚珠内にできて「胚嚢」、♂配偶体は花粉嚢ないし葯にできて「花粉粒」と呼ばれる。
種子植物の配偶体はさらに小さく、有性生殖器官(いわゆる「花」)の内部に作られる。♀配偶体は胚珠内にできて「胚嚢」、♂配偶体は花粉嚢ないし葯にできて「花粉粒」と呼ばれる。
7-1-4. 花粉粒=種子植物の雄性配偶体

右図は被子植物の小胞子・雄性配偶体形成の模式図だ。
-
小胞子母細胞(1)から減数分裂(rd)で小胞子4個の塊=四分子(2)ができる
-
四分子がばらけ、各々の小胞子(3)が成熟(m)して2細胞または3細胞の雄性配偶体=花粉粒(4)となる。葯壁最内層のタペート組織は、花粉成熟に必要な養分を供給し、花粉粒を覆う花粉壁を付加する。
-
柱頭上で発芽(g)した花粉粒(5)から伸びる花粉管は、花粉管核と2個の精細胞を含んでいる
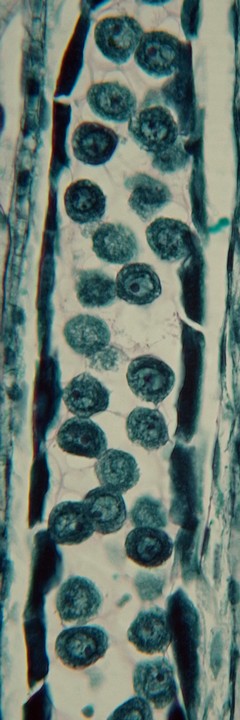
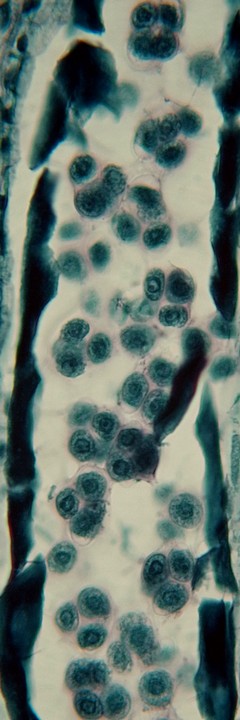
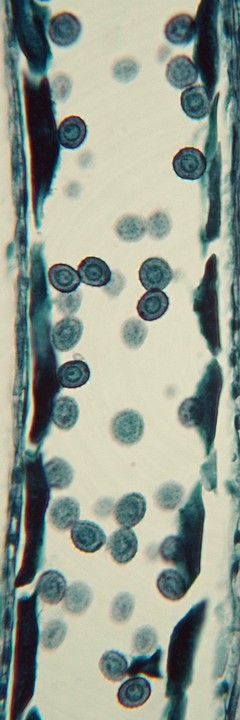
 ナベワリ(ビャクブ科)の若い葯で見られた減数分裂。
ナベワリ(ビャクブ科)の若い葯で見られた減数分裂。
葯室内の小胞子母細胞が減数分裂してできた小胞子×4は1つの塊(四分子[tetrad])となっており、まだ小さいつぼみを輪切りにすると、葯の断面には多数の四分子が観察される。
単子葉植物では、ナベワリのように、四分子の4つの小胞子が平面的に配列することが多い(ヤマノイモ科のような例外もある)。これに対して双子葉植物では、シャク・オオキンケイギクのように小胞子が四面体状に配置することが多い。
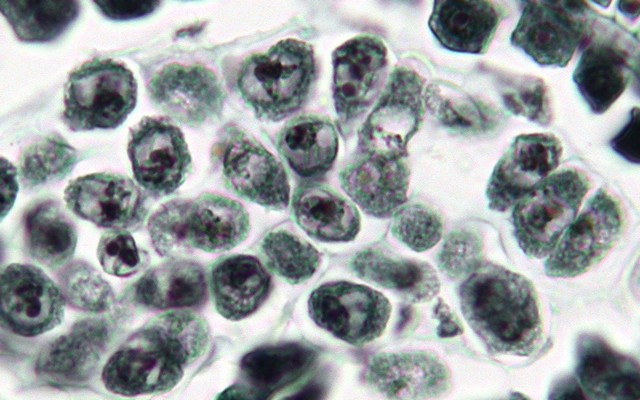

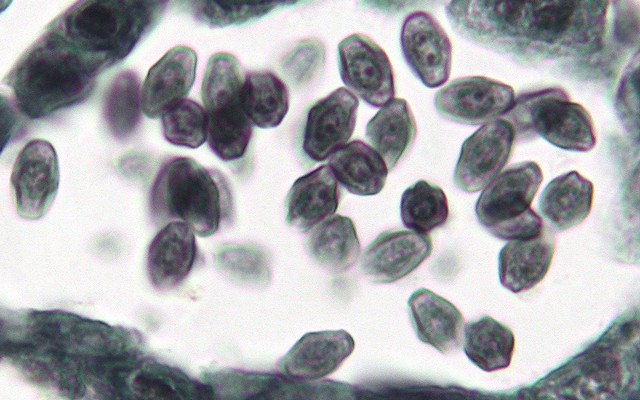
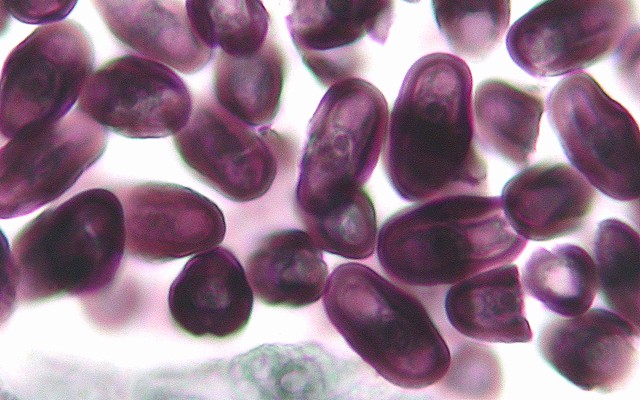
 上: シャク(セリ科)の花粉形成過程。花粉母細胞→減数分裂→未成熟な花粉→成熟した花粉。
上: シャク(セリ科)の花粉形成過程。花粉母細胞→減数分裂→未成熟な花粉→成熟した花粉。
左: シャクの雄しべ横断面。花粉粒を取り囲む濃く染色された細胞層(葯壁の最内層に当たる)はタペート層[tapetum]といい、花粉形成の各段階で大きな役割を果たす。
 オオキンケイギク(キク科)の花序の一部(断面)。外側(画像では左側)から順に咲いていくので、花粉の発達の段階を順に見ることができる(下の画像)。順に、花粉母細胞形成→花粉母細胞→減数分裂→花粉(未成熟)。葯壁内面のタペート層は全ての段階で濃く染色されている。
オオキンケイギク(キク科)の花序の一部(断面)。外側(画像では左側)から順に咲いていくので、花粉の発達の段階を順に見ることができる(下の画像)。順に、花粉母細胞形成→花粉母細胞→減数分裂→花粉(未成熟)。葯壁内面のタペート層は全ての段階で濃く染色されている。

裸子植物では、葯に当たるもの(小胞子の入っている袋=小胞子嚢[microsporangium])が小さな葉についていて、その葉(小胞子葉[microsporophyll])が集まって房のようになっている。
時間がたつにつれ、四分子はばらばらになり、個々の小胞子は葯壁最内層のタペート層から供給される栄養で成熟し、堅固な外殻(花粉壁)で覆われるようになる。小胞子は殻の中で少しだけ細胞分裂をして、裸子植物では2~数細胞、被子植物では2細胞か3細胞(種類によって決まっている)の雄性配偶体になる。雄性配偶体は(雌性配偶体もそうだが)、一つの大きな細胞の中に他の細胞があるという変わった構造をしている。花粉壁で覆われた雄性配偶体を「花粉粒」と呼ぶ。


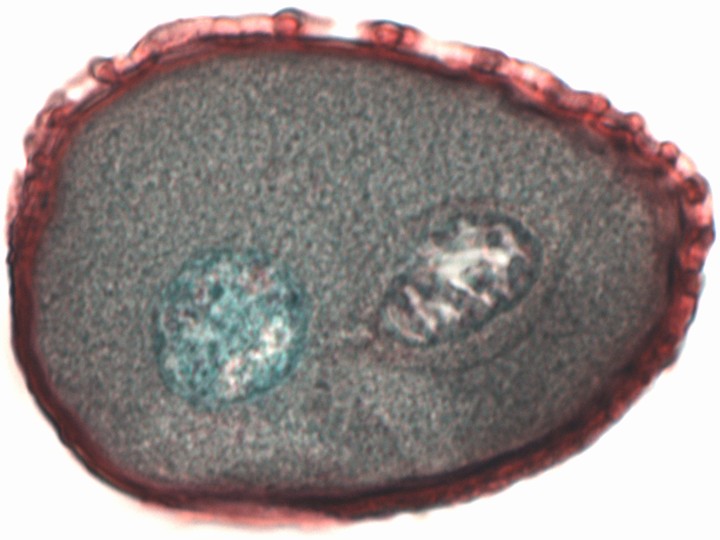
ウマノアシガタの成熟した葯の断面。花粉粒の中には2つの細胞核が見える。

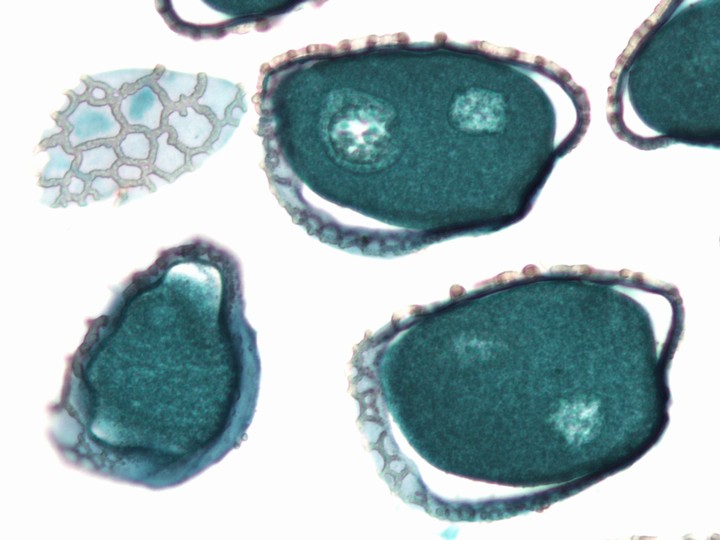
タカサゴユリ(ユリ科)の裂開した葯の断面と花粉粒。花粉粒の中に花粉管核と精細胞1個ずつが見える。
花粉粒は植物の種類によってさまざまな全形・サイズ・発芽口の特徴・表面パターンを示す(→花粉粒)。また、花粉壁はスポロポレニンという耐久性の高い物質でできていて、地層の中で分解されずに残る。このことを利用して、地層の中の花粉の組成を調べ、過去の植生を推定することができる。




上左・上右: シナレンギョウ(モクセイ科)の三溝粒
下左: コブシ(モクレン科)の単溝粒、下右: ヤマモモ(ヤマモモ科)の三孔粒
 ツツジ(ツツジ科)の花粉四分子。ツツジ類などでは小胞子の四分子がばらけないまま花粉四分子となり送粉されるため、成熟した花粉でも四分子の配列や発芽口の位置が分かる。
ツツジ(ツツジ科)の花粉四分子。ツツジ類などでは小胞子の四分子がばらけないまま花粉四分子となり送粉されるため、成熟した花粉でも四分子の配列や発芽口の位置が分かる。
7-1-5. 胚嚢=種子植物の雌性配偶体は胚珠に内蔵されている
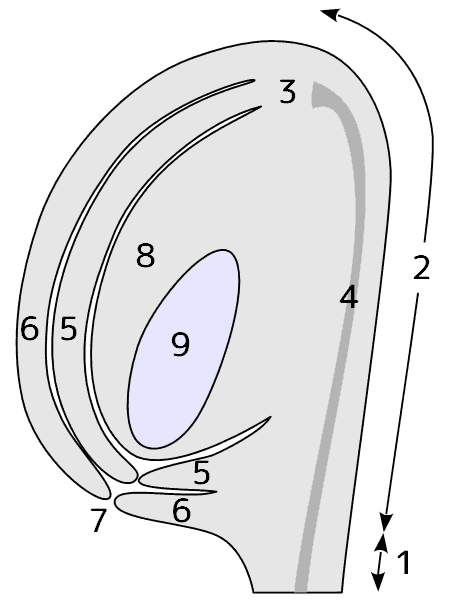
種子植物の胚珠は、珠心[nucellus]とそれを取り囲む珠皮[integument]からできている。珠心のまわりの組織が盛り上がり、ちょうどあんパンをつくるときにパン生地であんを包み込むようにして、珠心を包み込む。あんパンのへそと同じで、最後に包み込まれたところには、すきまが残る。このすきまを珠孔[micropyle]といい、(例外はあるが)受精のときに花粉管の入り口となる。
被子植物の胚珠の模式図。1―珠柄、2―ラフェ、3―カラザ、4―ラフェ維管束、5―内珠皮、6―外珠皮、7―珠孔、8―珠心、9―胚嚢
大胞子・雌性配偶体の形成には、さまざまなパターンがあるが、基本パターンでは、珠心の細胞の一つが減数分裂してできた4細胞のうち1個だけが大胞子で、細胞分裂して雌性配偶体になる。減数分裂でできた4細胞が、雄では4個とも、雌では1個だけが生殖に使われるのは多細胞動物と共通だ。
裸子植物・被子植物の一部では、減数分裂第2分裂の1つが省略され、3細胞しかできない場合がある。被子植物には、第2分裂で細胞壁が形成されず2核の大胞子ができるもの、第1・第2いずれも細胞壁が形成されず4核の大胞子ができるものがある。

被子植物の大胞子・雌性配偶体形成の例(模式図)。大胞子母細胞(1)から減数分裂で4細胞ができ、奥の1個が大胞子(2)となる。大胞子は、核の分裂による8核期(3)を経て8核7細胞の雌性配偶体=胚嚢(4)となる。
被子植物の雌性配偶体は、胚嚢 [embryo sac]と呼ばれ、巨大な中央細胞と中央細胞に埋め込まれた数細胞―珠孔近くの卵細胞と助細胞×2と反対側の0~3個の反足細胞―からなる。中央細胞は核を2個持っている(2個が融合して1個になっていることもある)。
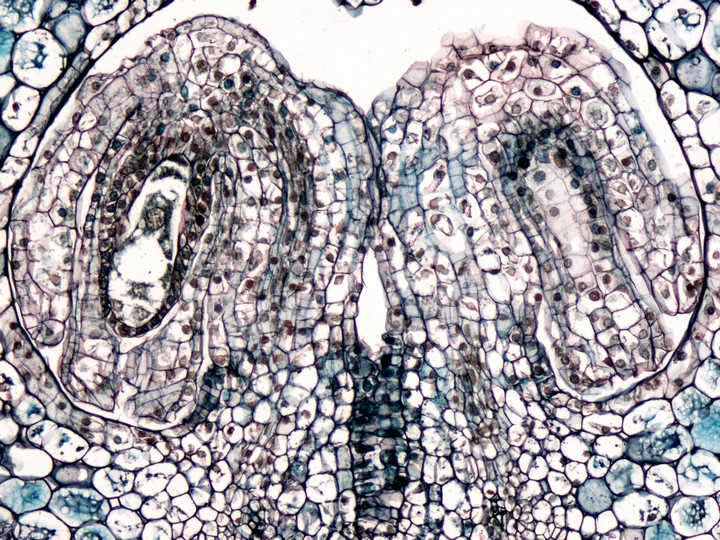 タカサゴユリ(ユリ科)の胚珠と胚嚢。胚珠の先端は下を向き、珠心を2枚の珠皮が二重に包んだ構造をとる。珠心の中央部は胚嚢で占められ、中央細胞の核(2個が融合して1個になっている)が中央に、先端部に卵細胞と助細胞×2がある。
タカサゴユリ(ユリ科)の胚珠と胚嚢。胚珠の先端は下を向き、珠心を2枚の珠皮が二重に包んだ構造をとる。珠心の中央部は胚嚢で占められ、中央細胞の核(2個が融合して1個になっている)が中央に、先端部に卵細胞と助細胞×2がある。

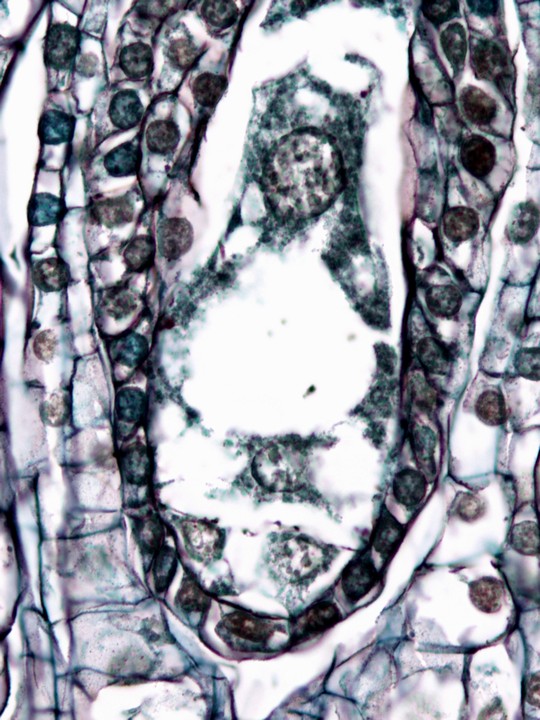
裸子植物の雌性配偶体は多数の細胞になって胚珠の大部分を占める。複数の造卵器(卵細胞とそれを囲む数細胞)ができ、残りの組織は受粉・受精の後も種子の貯蔵組織としてはたらく(ギンナンの食べる部分はほとんどが雌性配偶体だ)。
7-1-6. 胚珠と種子
-
「被子植物」=「胚珠(「子」)が覆われている植物」
-
「裸子植物」=「胚珠(「子」)が裸でいる植物」
名称の通り、被子植物の胚珠は、雌しべの中に入っていて、外界からは隔てられている。裸子植物でも、胚珠が外から丸見えというよりはある程度は隠されているものが多いが、雌しべはもっと完全に胚珠を包み込んでいるので、花粉管は雌しべの柱頭や花柱の組織の間を通り抜けなくては珠孔に入れない。
イチョウでは胚珠の上半分が露出している。ソテツでは大胞子葉、球果類(マツやスギなど)では芽鱗と苞鱗に覆い隠されており、ある程度は外から保護されている。イヌマキは胚珠が裸でついているように見えるが、実際には胚珠の周りを薄皮が覆い、珠孔だけが袋の口から顔を覗かせている。
珠孔に花粉管の先が入ると、そこから2個の精細胞が胚嚢に入り、そのうち1個が卵と融合する。もう1個は中央細胞の核と融合する。受精卵は細胞分裂をして胚に発達し、受精した中央細胞は細胞分裂をして内乳(内胚乳)[endosperm]となり、胚乳(種子の貯蔵組織)としてはたらく。このように受精が2カ所で起きること(重複受精[double fertilization])、受精でできた2つの細胞が両方とも組織に発達するのは被子植物の特徴だ。裸子植物にの一部でも重複受精が見られるが、受精卵の方だけが組織に発達する。
珠心の組織が胚乳となる場合や(スイレン・アカザなど; このような胚乳を「外乳」(外胚乳)[exosperm]と呼ぶ)、あるいは子葉が貯蔵の役割をするもの(キク科・マメ科など)もかなり多い。
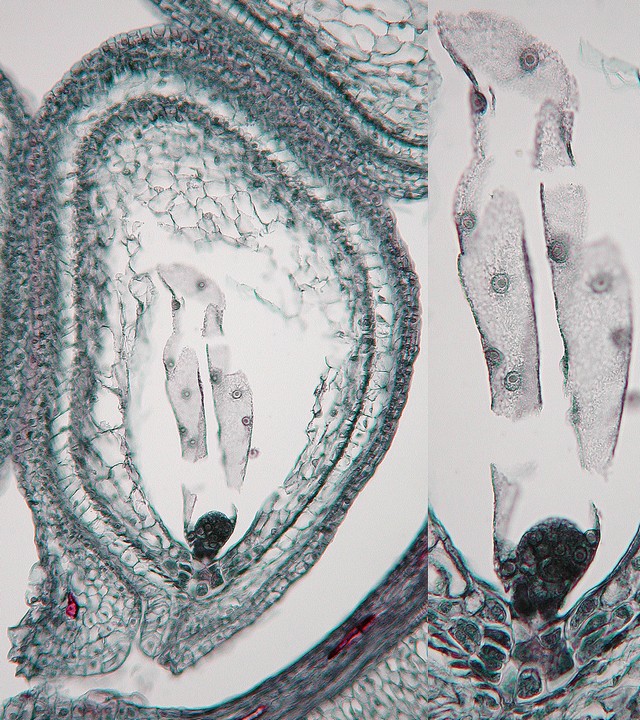
ナガバタチツボスミレ(スミレ科)の受精後の胚珠。珠孔近くの黒い塊状のものが胚、その上の大きな多核細胞が内乳だ。被子植物の内乳は、このようにまず核だけが続けて分裂し、ある程度核の数が多くなったあとで核と核との間に細胞壁ができることが多い。
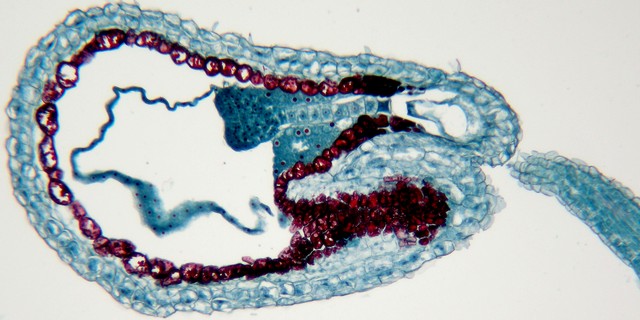
ナズナ(アブラナ科)の成長途中の種子の縦断面
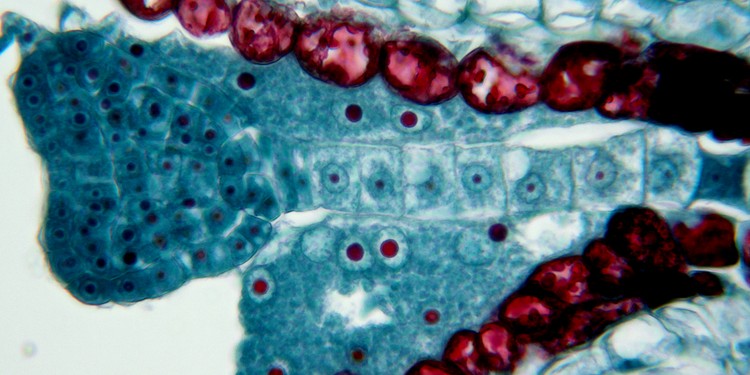
 上の図の一部の拡大。ハート形の胚と胚につながる細胞列(胚柄)は、ともに受精卵から発達する。多数の核を含む内乳は、ナズナの場合、種子が熟するまでに消えてしまう。
上の図の一部の拡大。ハート形の胚と胚につながる細胞列(胚柄)は、ともに受精卵から発達する。多数の核を含む内乳は、ナズナの場合、種子が熟するまでに消えてしまう。
種子は全体を種皮におおわれているが、胚珠から引き継いだ2つの構造、珠孔と珠柄の取れた痕(へそ[hilum])が残っている。大多数の種類では、珠孔とへそは隣り合っている。




オニバス(スイレン科)の種子。ほぼ球形で、深いオリーブ色の堅く厚い種皮が胚と胚乳を包む。へりにへそ(左側の楕円形)と珠孔(右側の円形)がある
種子の大きさ・形、また、表面のパターン(細胞の形・色・突起)は、きわめて多様で、種類の判別に役立つことも多い。
テキスト目次に戻る
ホームに戻る
