4-6. 食べられるのを防ぐしくみ
「食べられる」ことは、ほぼ全ての植物が直面し、生存と繁殖に大きく影響する現象だ。植物、特に葉や茎の特徴の中には、「食べられること」を防ぐため、あるいは被害を少なくすると思われるものが多い(前で述べたように、秋の紅葉もアブラムシの食害を避ける利点があるという有力な説がある)。
4-6-1. さまざまな生物がさまざまな方法で植物を「食べる」
生物が他の生物を食べることを食害/捕食/摂食 [predation]、食べられる(食害を受ける/捕食される/摂食される)ことを被食という。
食べる側をプリデーター(プレデター/食害者/摂食者)[predator]という。植物を食べる摂食者を植食者[phytophagous predator]といい、草食者/葉食者[hervivore]・果実食者[frugivore]・種子食者などに細分される。
虫媒花にとっての蜜や、動物散布果実にとっての果肉のように、食べられることが植物にとって利益となり、摂食であっても食害でない場合もあるので、摂食の方が幾分中立的な表現だ。捕食は、食べられるのが動物の場合に使われ、植物では滅多に使わない。
内部や表面に住みついて栄養を奪うことを寄生 [parasitism]といい、奪う方の生物を寄生者(パラサイト) [parasite]、奪われる生物を宿主(ホスト/寄主) [host]という。寄生による植物の被害を病害(植物病害)といい、病害を起こすパラサイトを病原体[pathogen]、病原体の寄生が始まることを感染[infection]という。
寄生は広い意味での食害に含まれるが、ふつうは上のように区別される。しかし、両者の境目ははっきりしない。特に昆虫が植物を「食べる」場合は、食害・寄生のどちらでも良さそうなものも多く、しばしばまとめて「虫害」と呼ばれる。
「食べる―食べられる」の関係は、摂食者が属する生物群の分類学的多様性、摂食が行なわれるスケールの大小、摂食のスタイルなど、多様性に富んでいる。従って、摂食に対抗する植物の特徴も非常に多様だ。
摂食者は脊椎動物からウイルスまでさまざまなグループに属する。パラサイトでない摂食者では、主なものだけでも、脊椎動物、昆虫や陸生巻貝などが挙げられる。パラサイトには、菌類・バクテリア・ウイルス・原生生物(特に卵菌類)などの微生物、昆虫・ダニ・センチュウなどの小型動物が含まれる。
食害のスケールという点から見ると、シカの食害のように森林や草原の風景を一変させるスケールの大きなものから、潜葉虫のように1枚の葉の中で完結するものもある。
河川敷で草を食むヤギ
ハモグリバエ類などの潜葉虫は葉に潜り込んで柵状組織と海綿状組織を食べながら移動する
「食べ方」も千差万別で、全体を選り好みをせずに食べる動物もいれば、セルロースの少ない若い葉や軟らかい葉を好んで食べる動物、葉をまるごとではなく葉脈や表皮の堅い部分を避けて食べる動物もいる。また、セミやカメムシ、アリマキは、口吻(筒になったくちばし)を突き刺して篩管から液体を吸収する。
マグワ(クワ科)の葉を食べるカイコ(カイコガ幼虫) 上: カラムシ(イラクサ科)の葉を食べるフクラスズメ幼虫 アオツヅラフジ(ツヅラフジ科)果実から吸汁するクチナガチョッキリ ゾウムシに産卵され幼虫の食物となったスダジイ(ブナ科)の果実。落下後しばらくすると幼虫が穴を開けて出て来る。
植物を食べるにあたり、植物体の多くを占めるセルロースの消化に体内や体外の共生微生物を使う植食者は多く、個体数も莫大で、生態系の物質循環でも大きな比重を占めることが多い。反芻動物(シカやウシなど; 共生者はルーメン内微生物)・シロアリ(消化管内微生物または菌園の菌類)・ハキリアリ(菌園の菌類)などが挙げられる。
寄生性の微生物(菌類・バクテリア・ウイルスなど)では内部への侵入過程に多様性が見られる。菌類の一部では、植物表面の水滴や泥などに付着した胞子が発芽し、菌糸が特殊なしくみによってクチクラと細胞壁を貫通して入り込む。クチクラ・細胞壁を破ることが出来ない多くの微生物は、気孔や、昆虫の食害等によって出来た傷・穴から侵入する。ウイルスには、アブラムシのような吸汁昆虫が媒介者となるものも多く、口吻によって篩管内へと送り込まれる。植物表面の撥水性や異物に反応した気孔閉鎖は、微生物の侵入に対抗するしくみだ。微生物の侵入に対しては、細胞レベル・分子レベルのさまざまな対抗手段が知られており、植物病理学の分野で活発な研究が行なわれている。
Phragmidium属(?)のサビ菌に寄生されたオヘビイチゴ(バラ科) 黒穂病菌に感染したハナタデ(タデ科・左)とマコモ(イネ科・右)。感染したハナタデは穂が黒い胞子で充満する。寄生されたマコモの茎は肥大し、マコモダケ(マコモタケ)と呼ばれて食用になる。 ウイルスに感染して葉が黄変したヒヨドリバナ(キク科)。維管束に沿って感染が進む。
寄生による形態の変化
寄生によって、宿主の形態にはさまざまな変化が起こる。
衰弱: 養分収奪の直接の結果として起こる、葉の黄変やシュートの萎凋
宿主側による防御反応
寄生操作(寄生者による宿主操作)[parasite manipulation]: 寄生者が宿主の特徴を自らに有利な方向に変形する。植物の寄生者の中には、植物ホルモンなどを通じて植物の形態に変化を起こすものがいる。変化によって、寄生者は養分収奪や他宿主への分散の効率化や天敵からの防御などの利益を受ける。
ゴール
寄生操作の例として最もしばしば見られるのは葉・茎・芽の組織の異常肥大(ゴール[gall])で、昆虫の寄生による場合は虫こぶ(虫癭/虫えい ちゅうえい)と呼ばれる。多くの場合、異常成長した組織は肉厚になり、空洞や巻き込みによって外部から遮断された空間が形成され、天敵から守られ十分な餌に囲まれた寄生者のすみかとなる。
タブノキ(クスノキ科)の葉裏にできたタマバエの虫こぶ テリハノイバラ(バラ科)の葉にできたバラハタマバチによる虫こぶ タマバエ[gall midge]によってカラスウリ(ウリ科)の茎が肥大した虫こぶ(クキフクレフシ) アブラムシによってエゴノキ(エゴノキ科)の芽が肥大した虫こぶ
ウイルス・バクテリアから菌類・卵菌類・センチュウ・昆虫・ダニなど、きわめて多様な生物が植物にゴール形成を引き起こし、ゴールができる植物の部位も多岐にわたる。ゴールの定義を広く取れば、根粒や菌根のような相利共生の舞台となるものも含まれる。
Harris NO & Pitzschke A (2020) Plants make galls to accommodate foreigners:some are friends, most are foes. New Phytologist 225: 1852–1872 https://doi.org/10.1111/nph.16340
生物進化ではよくあることだが、虫こぶのようなきわめて有利な特徴に対しては、それを標的とする捕食者が進化する。有名な例として、セイタカアワダチソウ(キク科)は、原産地の北米では、茎の途中にタマバエの寄生による虫こぶができる。数種のハチや甲虫が、虫こぶの内部や表面に産卵してタマバエ幼虫や虫こぶの組織を幼虫の餌にする。また、アメリカコガラやセジロコゲラは、冬にタマバエ幼虫目当てに虫こぶを探し出す習性をもつ。
寄生去勢
寄生者が宿主植物の花序や花の形成を妨げることもある(寄生去勢[parasitic castration])。有性生殖に養分が費やされない分、寄生者が収奪できる養分が増えることになる。
ムラサキケマン(ケシ科)の葉裏に散らばるサビ胞子堆
キケマン属(ケシ科)では、サビ菌(菌類)に感染した茎は花序をつけなくなって有性生殖能力を失い、一方で葉が高い位置につくようになる。サビ菌にとって、前者は栄養の吸収に、後者は胞子散布に有利となる特徴だ。ファイトプラズマ(ゲノムサイズが非常に小さく細胞壁がないなどの特徴を持つバクテリアのグループ)に寄生されたヒメウズ(キンポウゲ科)の花茎も、同じようにひょろ長く直立して花の代わりに数枚の葉をつける。
ムラサキケマン(ケシ科)。左の2本は葉裏にサビ胞子堆が出ていて、右の2本は出ていない。前者は後者より葉の位置が高く、茎頂に花序ができない。 ファイトプラズマ(バクテリア)に感染したヒメウズ(キンポウゲ科)。非感染個体(左下)に比べて草丈が高くなり、花がつくところにの数枚の葉がつく。
動物における寄生操作
寄生操作のうち、動物の宿主の行動や形態が操られる場合は、古くから多数の例が知られており、新しい例の報告も相次いでいる。
ハリガネムシ(類線形動物)に寄生されたカマキリ・バッタ・コオロギが水に飛び込み、多くの場合溺死するか魚などの餌食となる。ハリガネムシは水中に移行して繁殖を行う。
Leucochloridium(吸虫)に寄生された陸貝が枝先など目立つ場所に移動し吸虫のいる触角部をイモムシのように動かす(次のホストである鳥に捕食されやすくなる)
Wolbachia(バクテリア)は節足動物に寄生してさまざまな生殖器官の変化を引き起こす。その一つでは、寄生された昆虫やダンゴムシの雄は生殖器官が雌化して雌として生殖を行う(Wolbachiaは卵細胞を通じて母子感染するが、父子感染は起こらない)。
寄生バチに寄生されたガの幼虫が、身体から出た寄生バチ幼虫が作った繭を保護する(Grosman & al. 2008)
寄生操作は、大変インパクトが強く知名度も高い現象だが、それだけでなく、多様な生態系やグループに普遍的に存在して進化や生態系に大きな影響を与えているのではないかと考えられている。
Grosman AH, Janssen A, de Brito EF, Cordeiro EG, Colares F, Fonseca JF, Lima ER, Pallini A, Sabelis MW (2008) Parasitoid increases survival of pupae by inducing hosts to fight predators. PLoS ONE 3(6): e2276. DOI: 10.1371/journal.pone.0002276. link
寄生操作する寄生者に寄生して寄生操作することを「超操作」[hypermanipulation]という。米国南東部に分布するカンオケバンニンバチは超操作の例で(Weinersmith & al. 2017)、カシ(ブナ科)の若枝に虫こぶを作るタマバチに寄生する。寄生されていないタマバチは羽化後に虫こぶの壁に穴を空けて外に出るが、寄生されたタマバチは小さ過ぎて脱出できない穴を空け、頭部が穴に嵌まり込んだ状態で死亡する。タマバチより小形のカンオケバンニンバチは、タマバチの頭部を食い破って穴から外へ出る。タマバチに「空けさせた」穴を使って虫こぶから脱出することで、カンオケバンニンバチの生存率は自力で穴を空ける場合の4倍になる。
Weinersmith KL, Liu SM, Forbes AA, Egan SP. 2017. Tales from the crypt: a parasitoid manipulates the behaviour of its parasite host. Proceedings of the Royal Society B. Royal Society. doi:10.1098/rspb.2016.2365. link
食害/寄生の多様性に対応して、食害/寄生を避けるしくみも、ミクロレベルのしくみ(ここでは扱わない)を含めて、きわめて多様だ。
4-6-2. 有毒/忌避物質を含有する化学的防御
植物は、グループや種類によって異なるさまざまな物質(二次代謝産物)を合成し、その中に食害を防ぐはたらきを持つものが含まれる。タンニンや蓚酸カルシウムのように食物としての質を下げるものから、アルカロイド・青酸・精油(匂いがある揮発性の有機化合物)などの「植物毒」と呼べるようなものまである。
タンニンと食害の関係は、森林で大量に落下するために生態的な重要性が高いブナ科の果実(ドングリ・クリなど)で多数の研究がある。濃度が低い果実が食害動物に好まれる例(ex. Smallwood & Peters 1986)、高濃度の果実を給餌すると生存率が下がる例(Shimada & Saitoh 2003)、濃度が高いことで菌類の侵入が妨げられる例(Takahashi et al. 2009)などが知られている。
Shimada T, Saitoh T (2003) Negative effects of acorns on the wood mouse Apodemus speciosus . Population Ecology 45(1): 7-17 DOI 10.1007/s10144-002-0134-4 Abstract
Smallwood PD, Peters WD (1986) Grey squirrel food preferences: the effect of tannin and fat concentration. Ecology 67:168-174. Abstract
Takahashi A, Ichihara Y, Isagi Y, Shimada T (2010) Effects of acorn tannin content on infection by the fungus Ciboria batschiana . Forest Pathology 40(2): 96-99. DOI: 10.1111/j.1439-0329.2009.00612.x Abstract
身近な毒草の例。スイセン(ヒガンバナ科)・アジサイ(ユキノシタ科|アジサイ科)、どちらも全草が有毒だ。
実害がある濃度の植物毒を含む植物は身近にもきわめて多く、特にアルカロイドによる食中毒はけっこうな頻度で起こっている。ジャガイモ(ナス科)のソラニン、イヌサフラン(ユリ科|イヌサフラン科)のコルヒチン、ヒガンバナ科のリコリン、トリカブトのアコニチンなど。クワズイモやマムシグサ類(ともにサトイモ科)では、葉柄やイモ、果実に含まれる多量のシュウ酸カルシウム結晶が口の中や消化管にダメージを与えて食中毒になることがある。
クスノキ(クスノキ科)の葉の断面(パラフィン切片像・サフラニン―ヘマトキシリン―ファストグリーン三重染色)。柵状組織や海綿状組織にある大きな空洞は、精油を貯える大型の細胞で、「精油細胞」と呼ばれる。クスノキの精油は防虫作用がある樟脳(しょうのう)の原料となる。
植物毒を「毒」「薬」として使う人間/動物
ハシリドコロ(左)とタンナトリカブト(右)
人間は古くから植物毒を「毒」として狩猟・漁獲や戦闘に使ってきた。また、用法・用量を調節することで「薬」として用いる例も無数にある。ジギタリス(ゴマノハグサ科|オオバコ科)の毒性成分は強心剤として、ベラドンナやハシリドコロ(ともにナス科)の毒性成分(アトロピン)は散瞳剤や麻酔薬などに、古くから使われてきた。トリカブト類(キンポウゲ科)の地下茎は生薬「附子」(ぶし)として用いられる。
動物にも、植物毒を「毒」または「薬」として使うものがいる。天敵から身を守る「毒」として使う動物は、昆虫で多数の例が知られている。ジャコウアゲハ類(食草ウマノスズクサ科ウマノスズクサ属: アルカロイドのアリストロキア酸を含有)やマダラチョウ類(食草キョウチクトウ科ガガイモ類: アルカロイドを含有)は、食草に含まれる毒を身体に貯える(たいていは、派手な色彩と組み合わさっている)。マダラチョウは、成虫もピロリジジンアルカロイド(PA)を微量含む蜜を出す花から好んで吸蜜し、PAを天敵からの防御と性フェロモンの両方に使う。哺乳類では、アフリカタテガミネズミが毒のある樹皮を噛み砕いて有毒成分の溶け込んだ唾液を体側の毛にしみこませる(Kingdonら 2011)。
ウマノスズクサ上のジャコウアゲハ幼虫とブルーベリーから吸蜜するジャコウアゲハ成虫
「薬」として使う動物も報告されている。毒性がある植物を積極的に食べて体内の寄生虫を駆除する行動は、多数の哺乳類やヒトリガ類の幼虫(毛虫)で知られており、チンパンジー・ボノボ・ゴリラで最も研究が進んでいる(Huffman 2003, 2012; ハフマンら 2000; Singerら 2009)。
Kingdon J & al. (2011) A poisonous surprise under the coat of the African crested rat. Proc. R. Soc. B 279(1729): 675-680 link
Huffman MA (2012) 霊長類の自己治療行動―予防と治療(PDF) (神戸アニマルケア国際会議2012)
マイケル・A・ハフマン/小清水弘一/大東肇. 2000. サルの薬膳料理. 『霊長類生態学: 環境と行動のダイナミズム 』(京都大学出版会) 第4章 PDF
Huffman MA (2003) Animal self-medication and ethno-medicine: exploration and exploitation of the medicinal properties of plants. Proceedings of the Nutrition Society 62(2): 371-381. DOI: 10.1079/PNS2003257 link
Singer MS, Mace KC, Bernays EA (2009) Self-Medication as Adaptive Plasticity: Increased Ingestion of Plant Toxins by Parasitized Caterpillars. PLoS ONE 4(3): e4796. doi:10.1371/journal.pone.0004796 link
食害や感染による傷害が起こった時点で発動する化学的防御もさまざまな植物で知られている。傷害を受けた植物体ではジャスモン酸(JA)がすばやく生合成され、タンパク分解を妨げる物質(食害者の消化を妨げ、食物としての質を下げる)や感染抵抗性を持つ物質の合成など、多面的な防御反応を引き起こす。また、メチル化して揮発性のジャスモン酸メチル(MeJA)となり、周囲に発散して隣接する植物の防御反応を誘発するとともに、食害者の天敵を引き寄せる。
ジャスモン酸は植物ホルモンとして傷害応答以外にも多様な機能を持つ。ジャスモン酸メチルはジャスミン(モクセイ科)などの花の香りの主要な成分だ。
4-6-3. トゲや毛、粘液による物理的防御
トゲは大型の動物に、毛と粘着帯は小型の動物に対して効き目が大きい。
トゲを持つ樹木には、背が低いときにはたくさんのトゲをつけるが、成長するにつれトゲの数が少なくなる(またはつけなくなる)ものが多い。
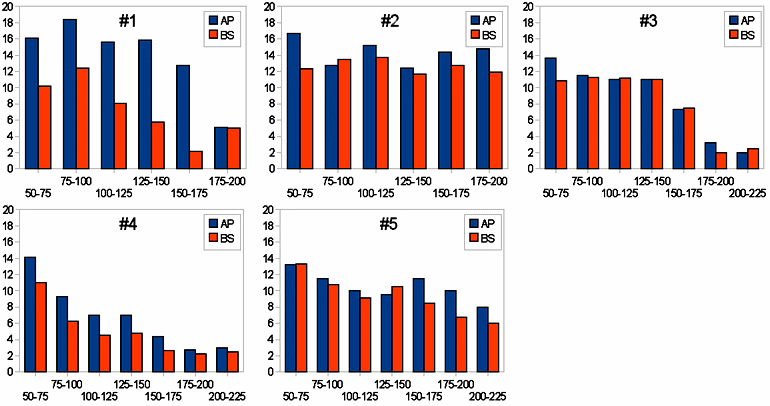
ヒイラギモクセイ(モクセイ科)の葉の縁にはギザギザ(鋸歯)があり、鋸歯の先端は鋭いトゲになっている。写真では、1本の木から3枚の葉を選んでいる。地面に近い枝ではトゲの数が多く、高いところにある枝ではトゲが少ない葉やトゲがまったくない葉が多い。 大学内に植えられている5本のヒイラギモクセイで計測した、葉の高さ(地表面から)とトゲの数の関係。各値は8~10枚の葉の平均値で、"AP"は枝の先端にある葉、"BS"は枝の基部にある葉。 ヒイラギ(モクセイ科)の枝。葉の縁には、先がトゲになった鋸歯がある。低い枝や小さな株では鋸歯は鋭く、トゲも長い。高い枝では、鋸歯の尖り方は緩くなり、トゲも短い。枝のつけねの方では鋸歯の数が少なくなり、中には全くない葉も出てくる。ヒイラギモクセイは交雑でできた園芸種で、トゲの性質は片方の親と推定されているヒイラギから受け継いだものと考えられる。 カラスザンショウ(ミカン科)。羽状複葉を持つ。若木のときには葉軸や枝に棘が多いが、アゲハの食害にはなすすべがない。 大きな木ではほとんど棘がなくなる。
イネ科のように細長い葉をもつ植物では葉縁に鋭いトゲが並ぶ例が多い(鋸歯と見なすこともある)。トゲの先端が葉の先端方向を向く場合が多く、葉の基部へと進む食害者に対する抵抗となる。イネ科では、ガラス質で補強されたトゲによって、葉の基部に囲まれた茎頂と新葉が守られる。
セイヨウヒイラギ(モチノキ科)では、草食動物に葉がついばまれた個体はトゲのある葉ができやすくなる。トゲのある葉とトゲのない葉の間でDNAメチル化の程度に違い(トゲのある葉の方が低い)が検出された。
Herrera CM, Bazaga P. (2012) Epigenetic correlates of plant phenotypic plasticity: DNA methylation differs between prickly and nonprickly leaves in heterophyllous Ilex aquifolium (Aquifoliaceae) trees. Botanical Journal of the Linnean Society. DOI: 10.1111/boj.12007
link ニュース記事
トゲには、茎や葉に当たるもの、葉の一部が変化したもの、表皮の突起に由来するものなど、さまざまなものがある。また、食害回避よりも他の植物に絡みつくことが主な役割と考えられるトゲも少なくない。 →
トゲ
成長中の若くやわらかい葉では毛が密についていて、成長が進んで硬くなるにつれて取れてしまうことが多い。
シロダモ(クスノキ科)の若葉は毛に覆われて白~金色に輝くが、成長するにつれてほとんどの毛が取れてしまう シロダモ(クスノキ科)の葉の表面。左から、新葉・やや成長した新葉・古い葉、上が葉の上面(向軸面)・下が下面(背軸面)。
植物体表面の粘液は、昆虫のような小形食害者の動きを妨げる。ムシトリナデシコ(ナデシコ科)では、茎の一部が粘液が滲み出す粘着帯となっている。ノアザミ(キク科)では花序を囲む総苞片が粘液を出す。腺毛によって弱い粘着性を帯びた茎や葉は、多数の植物で見られる。
上: ムシトリナデシコ(ナデシコ科)の花の下の茎は、一部が茶色い粘着帯となっている クサイチゴ(バラ科)の葉柄・葉軸には白い細毛と赤い腺毛の2種類がある
枝や葉が損傷を受けた断面から、テルペン類などの揮発性成分を含む粘液(樹脂/レジン[resin])が滲み出る種類もある。樹脂は昆虫や微生物を物理的に拘束するだけでなく、化学的防御に関わる物質を含む。典型的な樹脂は透明で黄色~褐色を帯びており、揮発性成分がなくなると固まって「ロジン[rosin]」と呼ばれる。
エンジュの大枝の切口から染み出した粘液が凝固している
粘液状態・固化状態とも樹脂/レジンと総称することもある。また、粘液状態のレジンのうち、有用性が高いものの一部は「バルサム[balsam]」と呼ばれる。
クロマツ(マツ科)の枝の切口
裸子植物球果類は樹脂の量が多く、特に、マツ属から採取される松脂(松ヤニ[pine resin])は、揮発性成分(松テレピン油[pine turpentine])が溶剤などに使われ、ロジンの方は粘性・可塑性を生かした多様な用途に使われる。そのため、単に「ロジン」と言うときは松脂のロジンを指すことが多い。
4-6-4. 天敵を常駐させる生物的防御
アリを招く花外蜜腺
イタドリ・アカメガシワ・サクラ・ホウセンカ・ヘチマなどでは、葉柄や葉身のつけねにイボ状の突起やクレーター状の丸いくぼみがある。これらの突起やくぼみは、特に若葉や若枝では、蜜を分泌し、花外蜜腺[extrafloral nectary]と呼ばれる。
フヨウやオクラ(アオイ科)では、萼片のつけね向軸面から蜜が出る。花の外側に分泌されるので、広い意味では花外蜜腺に含めることができる。
花外蜜腺には蜜を集めるアリが見られることが多い。また、アリの種によっては、他の巣のアリに奪われないように蜜腺に常駐することもある。葉の基部にアリがいることは、食害昆虫の移動や摂食を妨げると考えられている。
→ 花外蜜腺
ソメイヨシノ(バラ科)。サクラの仲間は、葉身基部か葉柄に蜜腺がある。 オクラ(アオイ科)。萼の基部からしみでる蜜にアリが集まっている
ナス科ナス属のSolanum dulcamaraは花外蜜腺は持たないが、食害による傷口から糖分の多い液を分泌してアリを誘引する。
植物を守る甘い出液 (Nature Plants: 注目のハイライト)
ダニ室
樹木の葉の裏面の葉脈分岐点には、小さな空洞(クスノキなど)・ポケット状の構造(ホルトノキなど)・細かい毛の茂み(ヤマボウシなど)など、微細な生物の「隠れ家」になるような構造(ドマティア[domatia])がある。これらにはダニの住居・産卵場所として使われることが多いので、「ダニ室」と呼ばれる。ダニ室には食害をするダニがいることもあるが、他のダニを捕食するダニが住んでいることもある。
クスノキ(クスノキ科)の葉の基部。脈の分かれ目のところが膨らんでいる。裏側には小さな穴があり、中にダニが見える。 ホルトノキ(ホルトノキ科)の葉脈分岐点にはポケットがある。若い葉のポケットには、しばしばカブリダニ(肉食性のダニ)が見られる。 ヤマボウシ(ミズキ科)の葉脈分岐点には茶色の毛がかたまっている。こういうところもダニのすみかとなる。 サンシュユ(ミズキ科)の葉脈分岐点。楕円形の暗い影はダニ。
4-6-5. 食害に対抗するさまざまな特徴
地表への退避
草食性哺乳類が多い場所では、低く地表付近に広がる草形が見られる種もある。
ツクシアザミ(キク科)。ふつうは左のように背が高くなる。シカの個体数が多いところでは、下のような低い円錐型の草姿になりながらも、鋭いトゲで茎頂を守り枯死を免れる。他の植物が減ったぶん株数は多くなる。 シカの食害圧下のバライチゴ(バラ科)
全草でなくても、分裂組織が地表すれすれの低いところにあれば、食害を受けても分裂組織が残って再生できる可能性が大きい。イネ科草本では、茎の頂端分裂組織・葉の介在分裂組織とも地表にあることが多く、草刈後にいち早く再生する様子が見られる。アフリカのサバンナ[savanna]や放牧地など草食性哺乳類が多い場所では、強い食害圧によってイネ科が優占する草原が広がる。
草刈りが行われる土手に広がるチガヤ(イネ科)の草原
同じ理由で、ひんぱんな草刈りが行われる場所も、しばしばイネ科の草原となる。
突発的な集中開花・結実
花や果実の数は、気候や日当たりの変化によって同じ個体であっても年毎に変わる。しかし、中には、一見説明がつかないほどの極端な変動をする植物がある。
左―開花しているネザサ(イネ科)・右―枯れ上がった桿が目立つ開花後のネザサ群落
タケ・ササ(イネ科)[bamboo]の多くは、数十年の寿命をもち、種によって決まっているとされる(マダケなどでは120年に及ぶといわれる)。寿命が来るまで開花せずに成長を続け、最期の年に地下茎に貯えた栄養の全てを費やして開花・結実して枯死する。1つ1つの笹原や竹藪は、数十年に一度、全てが一斉に開花・結実して大量の種子を生産し、枯れてしまう。
ブナやナラ(落葉性のコナラ属)では、2年から数年に一度、多くの個体が多数の種子をつける「豊作年」がある。豊作年の間に凶作年(多くの個体が種子をあまりつけない)と並作年(豊作年と凶作年の中間)が入る。このような、個体どうしが同調して豊凶の繰り返しことをマスティング[masting; mast seeding]という。
東南アジアの熱帯雨林では、1年から数年ごとに多数の樹種が種ごとに時期をずらしながら一斉に開花する現象が知られている(竹内 2014)。
毎年ある程度コンスタントに開花することと比べて、年毎に開花数が極端に変動することにどのような利点があるのかには、さまざまな説明があるが、主要なものは以下の2つだ(Kelly 1994, Kelly & Sork 2002)。
種子食害者の飽和
風媒による送受粉の成功率を高める
毎年n個の花が咲く
3年に1回3n個の花が咲く
を比べたとき、(B)では空中の花粉の密度が(A)の3倍で、受粉率(柱頭に花粉が1個以上つく確率)は、(A)の受粉率が0.1のときは(B)では0.271、0.2なら0.488となる。
これらの仮説は、大量開花の年とそうでない年で種子の食害率や結実率(花が果実となる割合)を比較することで検証できる。しかし、簡単に想像できるように、膨大な時間・労力か大変な幸運(またはその両方)を必要とする。
Janzen DH. 1976. Why bamboos wait so long to flower. Annul. Rev. Ecol. Syst. 7:347-391
Kelly D. 1994. The evolutionary ecology of mast seeding. Trends Ecol. Evol. 9(12):465-470. pdf
Kelly D, Sork VL. 2002. Mast seeding in perennial plants: why, how, where? Annu. Rev. Ecol. Syst. 33:427-447.
竹内やよい. 2014. ボルネオ熱帯雨林の一斉開花の要因を探る. 生命誌ジャーナル 81 link
テキスト目次に戻る ホームに戻る


 嗜好植物のアオキ(ミズキ科|ガリア科)。葉は対生し、大型で光沢が強く、粗い鋸歯がある。若い枝は深緑色。
嗜好植物のアオキ(ミズキ科|ガリア科)。葉は対生し、大型で光沢が強く、粗い鋸歯がある。若い枝は深緑色。


 シカが繰り返して葉を食べたことによって枯死したスズタケのやぶ
シカが繰り返して葉を食べたことによって枯死したスズタケのやぶ

 ヒカゲノカズラに埋め尽くされた林床(宮崎県えびの高原)
ヒカゲノカズラに埋め尽くされた林床(宮崎県えびの高原)

 ハモグリバエ類などの潜葉虫は葉に潜り込んで柵状組織と海綿状組織を食べながら移動する
ハモグリバエ類などの潜葉虫は葉に潜り込んで柵状組織と海綿状組織を食べながら移動する

 上: カラムシ(イラクサ科)の葉を食べるフクラスズメ幼虫
上: カラムシ(イラクサ科)の葉を食べるフクラスズメ幼虫



 Phragmidium属(?)のサビ菌に寄生されたオヘビイチゴ(バラ科)
Phragmidium属(?)のサビ菌に寄生されたオヘビイチゴ(バラ科)









 アブラムシによってエゴノキ(エゴノキ科)の芽が肥大した虫こぶ
アブラムシによってエゴノキ(エゴノキ科)の芽が肥大した虫こぶ ムラサキケマン(ケシ科)の葉裏に散らばるサビ胞子堆
ムラサキケマン(ケシ科)の葉裏に散らばるサビ胞子堆

 ファイトプラズマ(バクテリア)に感染したヒメウズ(キンポウゲ科)。非感染個体(左下)に比べて草丈が高くなり、花がつくところにの数枚の葉がつく。
ファイトプラズマ(バクテリア)に感染したヒメウズ(キンポウゲ科)。非感染個体(左下)に比べて草丈が高くなり、花がつくところにの数枚の葉がつく。



 ハシリドコロ(左)とタンナトリカブト(右)
ハシリドコロ(左)とタンナトリカブト(右)








 カラスザンショウ(ミカン科)。羽状複葉を持つ。若木のときには葉軸や枝に棘が多いが、アゲハの食害にはなすすべがない。
カラスザンショウ(ミカン科)。羽状複葉を持つ。若木のときには葉軸や枝に棘が多いが、アゲハの食害にはなすすべがない。 大きな木ではほとんど棘がなくなる。
大きな木ではほとんど棘がなくなる。



 シロダモ(クスノキ科)の若葉は毛に覆われて白~金色に輝くが、成長するにつれてほとんどの毛が取れてしまう
シロダモ(クスノキ科)の若葉は毛に覆われて白~金色に輝くが、成長するにつれてほとんどの毛が取れてしまう


 上: ムシトリナデシコ(ナデシコ科)の花の下の茎は、一部が茶色い粘着帯となっている
上: ムシトリナデシコ(ナデシコ科)の花の下の茎は、一部が茶色い粘着帯となっている


 クロマツ(マツ科)の枝の切口
クロマツ(マツ科)の枝の切口


 ソメイヨシノ(バラ科)。サクラの仲間は、葉身基部か葉柄に蜜腺がある。
ソメイヨシノ(バラ科)。サクラの仲間は、葉身基部か葉柄に蜜腺がある。 オクラ(アオイ科)。萼の基部からしみでる蜜にアリが集まっている
オクラ(アオイ科)。萼の基部からしみでる蜜にアリが集まっている



 ヤマボウシ(ミズキ科)の葉脈分岐点には茶色の毛がかたまっている。こういうところもダニのすみかとなる。
ヤマボウシ(ミズキ科)の葉脈分岐点には茶色の毛がかたまっている。こういうところもダニのすみかとなる。 サンシュユ(ミズキ科)の葉脈分岐点。楕円形の暗い影はダニ。
サンシュユ(ミズキ科)の葉脈分岐点。楕円形の暗い影はダニ。

 シカの食害圧下のバライチゴ(バラ科)
シカの食害圧下のバライチゴ(バラ科)



