2. 種子と芽生え
2-1. 種子
種子[seed]は、胚・胚乳・種皮からできている。



カキノキ(カキノキ科)の種子と断面。茶色の種皮が半透明白色の貯蔵組織(胚乳)を包み、胚乳の中に棒状の胚軸と子葉2枚を持つ胚がある。


カキノキ(カキノキ科)の芽生え。種皮の中の空間は子葉が占め、胚乳は種皮の裏側に薄い層として残るだけだ。成長した子葉の間には本葉が見える。胚軸は種子の倍の長さに伸びて、基部から発根している。
胚[embryo]
受精卵が分裂して出来た幼体。基本的には胚軸[hypocotyl]とその一端についた子葉[cotyledon]でできている。
胚軸は、名前の通りに胚の中心軸で、シュート系と根茎の元手となる。胚軸の両端には分裂組織[meristem](分裂能力がある細胞の集まり)がある。2つの分裂組織のうち、子葉の側の茎頂分裂組織からは植物の生涯で最初のシュートができ、反対側の根端分裂組織から最初の根ができる。
子葉の基本的な役割は、胚乳から養分を吸収して胚軸に受け渡すことだ。カキノキの種子では、発芽までに子葉が成長して胚乳の養分を吸収し、体積の多くを占めていた胚乳が種子の裏に薄く残るだけになる。子葉から養分を供給された胚軸は伸長・肥大して種皮から出て根を下ろす。このように、他物から養分を吸収する器官を吸器[haustorium; 複 haustoria]という。追加の機能をもつ場合もあるが、全ての植物に見られるわけではない。
-
貯蔵器官: 胚乳からの養分吸収は種子成熟までに完了し(種子成熟時点で胚乳は消失)、吸収した養分を発芽までの期間貯蔵する。子葉が貯蔵器官として働く種子を「無胚乳種子」という。
-
光合成器官: 発芽後に地上に出て面積を増し、初期の光合成を担う。このような種子発芽のタイプを「地上子葉性」という。
-
保護器官: 単子葉植物では、胚軸先端の茎頂を子葉の基部が鞘のように取り巻いて保護する(子葉鞘)
胚軸や子葉は、まっすぐ伸びていることもあるが、種子に比べて胚が長い場合には、くるりと丸まっていたり、もっと複雑に畳み込まれていることもある。種類によっては、種子が散布された時点では胚軸と子葉の区別がはっきりしない細胞塊の状態で、土壌中で胚が形成される。
胚乳[albumen]
デンプン・脂肪・タンパクを高密度で含む細胞で出来た貯蔵組織[storage tissue]で、胚を取り囲んでいるときが多いが、ハコベやヒユのように胚が貯蔵組織に取り巻くようになっているときもある。
胚乳の元となる組織や細胞は植物によってさまざまで、被子植物では内乳(胚嚢の中心細胞+精核)または外乳/周乳(珠心)から形成され、裸子植物では配偶体の組織が胚乳としてはたらく。

フデリンドウ(リンドウ科)の種子断面。有胚乳種子の例。
胚乳は、子葉によって養分が吸収される過程で細胞が崩壊し、消失する。カキノキやフデリンドウでは、種子が熟して散布される段階では胚乳が残っており、発芽の過程で子葉が胚乳の養分を吸収して胚に受け渡す。このような「散布段階で胚乳がある種子」を胚乳種子または有胚乳種子[albuminous seed]という。
ダイコンの種子では、種子が熟する過程で子葉が胚乳の養分を吸収し、種子散布までには胚乳が消失して、養分を貯蔵する子葉が種子の中身の大部分を占めるようになる。このような「散布段階で胚乳がない(あるいは、ほんの少ししかない)種子」を無胚乳種子[exalbuminous seed]と呼ぶ。有胚乳種子と無胚乳種子の違いは、子葉が吸収器官としてはたらくタイミングの違いだ。マメ科・アブラナ科・ウリ科・ブナ科など身近な双子葉植物には無胚乳種子の方が多い印象がある。


 ダイコン(アブラナ科)の種子。無胚乳種子で、赤褐色の種皮が胚を包む。胚は棒状の胚軸と2枚の子葉(貯蔵の役割を果たす)からできている。
ダイコン(アブラナ科)の種子。無胚乳種子で、赤褐色の種皮が胚を包む。胚は棒状の胚軸と2枚の子葉(貯蔵の役割を果たす)からできている。

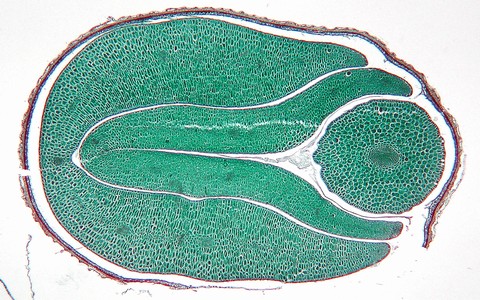 輪切りにしたダイコンの種子の断面。2枚の子葉と胚軸が折り重なるように納まっている。
輪切りにしたダイコンの種子の断面。2枚の子葉と胚軸が折り重なるように納まっている。
種皮[seed-coat]
胚と貯蔵組織をくるむ殻。胚珠の珠皮(1枚または2枚)の細胞層が、リグニンやタンニンなどの集積、細胞の拡大、細胞の崩壊などの変化を経て種皮を構成する細胞層となる。種皮の内部構造や表面の模様は、多くのグループで変化に富む。

ダイコンの種皮の断面構造。写真の上側が種子表面の方に当たる。珠皮に含まれる5つの細胞層がそれぞれ異なる特徴を持つ細胞層に変化して種皮となる。
硬い種皮は、さまざまな危険――動物や昆虫に食べられること、菌類や細菌に侵されること、衝撃や圧力で傷ついたり、ひしゃげたりすること――から中身を守っている。ただし、種類によっては、種皮は薄くて軟らかく、種皮を覆う果皮が主に保護の役割を担うものもある。
2-2. 種子の貯蔵養分

コムギ・イネ・トウモロコシ(イネ科)・ソバ(タデ科)・エンドウ・インゲンマメ・ダイズ・ラッカセイ(マメ科)・アーモンド(バラ科)・ゴマ(ゴマ科)・クリ(ブナ科)の種子の成分。左から水分(水色)・タンパク(濃赤)・脂質(淡黄)・炭水化物(灰)・ミネラル分(白)と並んでいる。炭水化物の多くはデンプンとして含まれる。同様の図とともに、日本食品標準成分表(文部科学省)による(食品成分データベース・日本食品標準成分表・資源に関する取組)。
貯蔵養分の組成は種類によって違う。炭水化物(糖質)を主体に貯め込んでいる種子を「デンプン種子(デンプン性種子)[starch seed]」といい、脂質を多めに貯め込んでいる種子を「油種子(脂肪性種子/油糧種子)[oil seed]」という。食用のデンプン種子は穀物と呼ばれる。

イネ科の種子の成分
イネ科(左図)やヒユ科(アマランサスなど)・ソバ(タデ科)などの穀物の種子は炭水化物の比率が高く、70~80%にのぼる。イネ科の食用種子はシリアル[cereal]と呼ばれる。

マメ科の種子の成分
根粒バクテリアとの共生によってタンパク合成に必要な窒素を得る能力が高いマメ科では、種子(マメ科の食用種子はパルス[pulse]と呼ばれる)に含まれるタンパクの比率が高い(左図)。栄養的に互いに補う関係にある穀物と豆類の組み合わせは、農耕の基盤となった。ダイズやナンキンマメ(ラッカセイ)はタンパクに加えて脂質の比率が高い油種子だ。

イネ科・マメ科以外のさまざまな科の種子の成分
種子が含む脂質は植物油[vegetable oil]の原料となる。マメ科の油種子からは大豆油・ピーナツオイルなど、また、左図の油種子からはアーモンドオイル・麻の実オイル・荏胡麻油・芥子油(ポピーシードオイル)・胡麻油・松の実オイルが作られる。他には、ベニバナ油(サフラワー油)・ヒマワリ油・菜種油(キャノーラ油)なども油種子由来だ。

イネ(イネ科)の種子(玄米)・胚(米糠)・胚と皮を取った種子(精白米)の成分
デンプン種子でも、トウモロコシではデンプン(コーンスターチ)を取った残り滓(コーンミール)からコーン油が、イネでは精白で出る米糠から米糠油が作られる。
種子以外では、アブラヤシ(ヤシ科)の果肉からヤシ油(パーム油)、オリーブ(モクセイ科)の果肉からオリーブオイルが作られる。世界での生産量は、大豆油とヤシ油が飛び抜けて多く、次いで菜種油、ヒマワリ油の順となる。アブラヤシの栽培は、東南アジアにおける森林破壊の主原因の1つだ。
現在では、植物油の多くは食用に使われる。昔は照明用にも植物油が多用されており、ずっとさまざまな植物の種子が油源として使われていた。最近ではバイオディーゼル原料としての利用が増えている。
バイオ燃料[biofuel]はエタノール[ethanol]とバイオディーゼル[biodiesel]がほとんどを占める。前者は穀物(特にトウモロコシ)やサトウキビなどから(糖化+)発酵で作られる。後者はヤシ油・大豆油・菜種油が主な原料で、脂質のグリセロールをメタノールに置換したものだ。
油種子は、炎で着火し、短時間だがろうそくの代わりになる(皮付きのままだと爆ぜることがあるので、皮を剥いた方が良い)。



ベニバナ(キク科)の果実に着火したところ



ダイズ(マメ科)の種子に着火したところ


トウモロコシ(イネ科)の種子は炙っても着火せずに焦げていく
2-3. 種子休眠と種子発芽
例外もあるが、種子は、水分・水溶性の養分・酵素の量が少なく(水分5~20%程度)、ほとんどの生命活動(代謝・生合成・細胞分裂)は停止に近い状態にある(種子休眠 [seed dormancy])。休眠状態の種子は、その代わり、普通だったら植物が耐えられないような条件(寒冷・高温・乾燥・暗黒)でも生き続け、長い寿命を保つことができる。
保管されていた種子や押し葉標本についていた種子が数十年~百年を経て発芽したという報告はまれではない。それをはるかに超えて、数千年~1万年前の種子(古代種子)が発芽したという報告もある。多くの場合、種子の年代は出土層や同時に出土した遺物の鑑定から間接的に推定される。1951年に千葉市で発掘された「大賀ハス」もその1つで、丸木舟の木片が放射性炭素年代測定で約3000年前と推定された。
直接測定した例では、中国東北部の湖跡から発掘されたハスの種子が放射性炭素年代測定で約1300年前と推定されている(Shen-Miller et al. 1995)。この報告では、発芽した種子を覆っていた果皮が年代測定に使われた。
-
Shen-Miller J, Mudgett MB, Schopf JW, Clarke S, Berger R (1995) Exceptional Seed Longevity and Robust Growth: Ancient Sacred Lotus from China. American Journal of Botany 82(11): 1367-1380. Article Stable URL: http://www.jstor.org/stable/2445863
条件が揃うと、休眠から醒め、種子はさまざまな生命活動を開始する。覚醒の条件は、植物の種類によって違うが、多くの場合、その植物の生育様式と対応している。例えば、寒い冬を避けて春に発芽する種子は、一定時間低温が続くと休眠から醒め、乾期が厳しい環境に生える植物の多くでは、土壌湿度によって休眠から醒める。対照的に、年間を通じて気温と降水が豊かな湿潤熱帯には休眠をほとんど行わないものが多い。また、他の植物との競合がない裸地や半裸地で成長する植物は、光と高温で休眠から醒める。


吸水してふくらんだダイズとアズキ(ともにマメ科)種子。それぞれ、左―吸水後・右―吸水前。
休眠から醒めた種子が最初に始めるのは、吸水・膨張と貯蔵物質の急速な代謝(分解してエネルギーや低分子[師管を通じて輸送されやすい]に変換する)だ。子葉から養分を供給された胚軸は、全体が伸びるとともに両端の分裂組織が細胞分裂を始め、幼芽と幼根が成長して、ふつうは先に幼根、ついで幼芽が種皮から突き出す(種子発芽[seed germination])。このようになった胚は「芽生え」[seedling](実生みしょう)と呼ばれる。
字面で「発芽」であっても、発根から始まる一連の過程を種子発芽といい、幼根の先端が顔を出したときから始まる。
食品としての芽生え
まだ光合成を始めていない、あるいは始めて間もない芽生えは、貯蔵された養分を代謝して分裂組織に送っている最中にある。
- 養分は貯蔵されているときより低分子となっており、消化吸収されやすい
- 代謝のために酵素や補酵素、その他さまざまな物質が活発に合成されている
- セルロースやリグニンの含量が低い
 ダイズ・リョクトウの種子ともやしの成分(水分を除く)。左からタンパク(濃赤)・脂質(淡黄)・炭水化物(灰)・ミネラル分(白)と並んでいる。日本食品標準成分表(文部科学省)による。
ダイズ・リョクトウの種子ともやしの成分(水分を除く)。左からタンパク(濃赤)・脂質(淡黄)・炭水化物(灰)・ミネラル分(白)と並んでいる。日本食品標準成分表(文部科学省)による。
ダイズ・リョクトウ(マメ科)で種子ともやしの成分を比較すると、もやしでは炭水化物と(ダイズでは)脂質の比率が減ってタンパクが増加している。また、ビタミンの含有量も増えており、KやCでは数十倍にのぼる。
 ダイズ・リョクトウの種子ともやしが含むビタミン類等の乾物中含有量。ダイズ種子の含有量を1とした相対値(ただし、ビタミンKとCは0.1)で示す。日本食品標準成分表(文部科学省)による。
ダイズ・リョクトウの種子ともやしが含むビタミン類等の乾物中含有量。ダイズ種子の含有量を1とした相対値(ただし、ビタミンKとCは0.1)で示す。日本食品標準成分表(文部科学省)による。

 リョクトウ(マメ科)もやし
リョクトウ(マメ科)もやし
これらは食品として有利な性質なので、さまざまな豆類の芽生えが「もやし」[bean sprout]として使われている。もやしのもう一つの利点は、貯蔵・運搬が容易な種子の状態からビタミンCをはじめとする野菜の養分を取り出せることで、かつては航海や軍隊でビタミンC欠乏症の予防に重宝された。現在店頭で普通に見られるものとしては、いわゆる普通のもやし(リョクトウ)、大豆もやし(ダイズ)、カイワレダイコン(ダイコン)や豆苗(エンドウ)などがある。麦芽[malt](オオムギの芽生え)では、貯蔵デンプンを分解するためにα-アミラーゼが多量に合成されており、糖の生産や酒造に使われる。
2-4. 芽生え
双子葉植物の芽生え

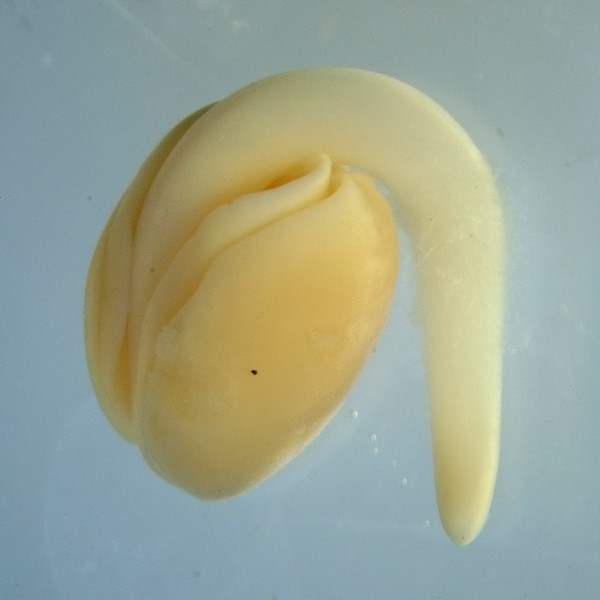
発芽直後のダイコン(アブラナ科)の芽生え。子葉は2枚が重なり、さらに胚軸を挟むように2つ折りになっている。
双子葉植物では、子葉は2枚(ごく少数だが1枚しかないものがある)で茎頂分裂組織は2枚の子葉の根元にはさまれた位置にある。

 |
 
子葉が展開したダイコンの芽生え。△▲は胚軸の両端を示す。2枚の子葉の葉柄にはさまれて、茎頂がある |

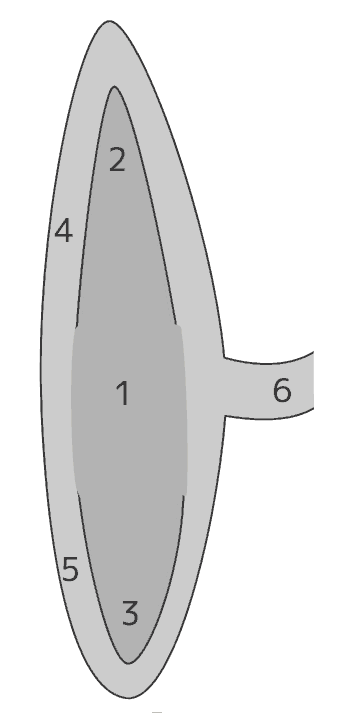
左の図は、双子葉植物の種子と芽生えのつくりを模式的に示している。芽生えでは、胚軸(5)の子葉側の端にシュート(幼芽+上胚軸)、反対側の端に両端に根(幼根)が1つずつあって、それぞれがシュート系と根系の出発点となる。
| 1 | 種皮 |
| 2 | 胚乳 |
| 3 | 胚 | 4 | 子葉[cotyledon] |
| 5 | 胚軸[hypocotyl] |
| 6 | 幼根[radicle] |
| 7 | 根端 |
| 8 | 側根 |
| 9 | 上胚軸[epicotyl] |
| 10 | 幼芽[plumule] |
| 11 | 茎頂 |
この図では、子葉と幼芽の間の茎を上胚軸としている。幼芽も含む子葉から上部、つまり胚がもつシュート全体を上胚軸と呼ぶ場合もある。
 双子葉植物の成長した植物体の模式図。
双子葉植物の成長した植物体の模式図。
H―胚軸[hypocotyl]、C―子葉[cotyledon]
シュート系
E―上胚軸[epicotyl]
N1―第1節、L1―第1葉(第1本葉)、S1―第1葉の腋芽から成長したシュート
N2―第2節、N2―第2葉(第2本葉)、S2―第2葉の腋芽から成長したシュート
N3―第3節、L3―第3葉(第3本葉)、B3―第3葉の腋芽
N4―第4節、L4―第4葉(第4本葉)、B4―第4葉の腋芽
BT―頂芽
根系
RT―主根
R1~6―側根
胚軸は茎とも根とも異なる維管束の配置を持つ。しかし、成長した植物体では、胚軸は茎の基部としてはたらくことが多く、日常生活上では茎の一部として扱われることがある。
双子葉植物の芽生えで、地上子葉性と地下子葉性の違いは、子葉が芽生えの光合成に大きく貢献するか、わずかしか~全く貢献しないかの違いだ。
双子葉・地上子葉性の芽生え



ホウセンカ(ツリフネソウ科)の芽生え。種皮を脱いでから、子葉は扁平になり、数倍の面積に成長する。
ダイコン(アブラナ科)・アサガオ(ヒルガオ科)・ヒマワリ(キク科)・ホウセンカ(ツリフネソウ科)・ワタ(アオイ科)などでは、子葉が種皮をかぶったまま地上に出てくる(地上子葉性/地上発芽)。種皮を脱いでから成長してより扁平で面積が広い形状になり、光合成をする。暗所では、カイワレダイコンのように、子葉の下の軸(胚軸)が伸長して子葉を高いところに持ち上げる。子葉柄が長い種類では、子葉柄が伸びたり、立ち上がる場合もある。子葉の寿命は相当長く、本葉が広がってからも、光が届く間は光合成が続く。子葉の間にある分裂組織から新しい葉が出てくるまでは、芽生えの生活は子葉が支えている。フラサバソウ(ゴマノハグサ科|オオバコ科)のように、開花・結実期まで子葉が残っている場合さえある。


|

|
|
フラサバソウ(オオバコ科)の芽生え。暗所(右)では胚軸が長くなり、子葉柄が直立する
|

フラサバソウの子葉は植物全体が枯れはじめるころまで残る
地上で展開する子葉は、普通葉(本葉)と形状が違うことが殆どで、さまざまな点で普通葉より特徴に乏しい単純な形状をしていることが多い。

|

|


|


|
|
子葉と本葉の形状の違い。カタバミ(カタバミ科・上段左)とクサイチゴ(バラ科・上段右)の本葉は複葉、イロハモミジ(ムクロジ科・中段左)とミヤマハハソ(アワブキ科・中段右)の葉は複雑に切れ込むが、子葉はこれらの特徴をもたない単純な形状をしている。アオカズラ(アワブキ科・下段左)のような左右非対称の腎臓形の子葉、ノグルミ(クルミ科・下段右)のように深く切れ込んだ子葉もあるが、少数派だ。
|
単子葉性双子葉植物
例外的に、芽生えに子葉が1枚しかない双子葉植物もある。ケシ科・キンポウゲ科・サクラソウ科・タヌキモ科・キク科などさまざまなグループで散発的に見られる。


ジロボウエンゴサク(ケシ科)の芽生えは子葉1枚で、後から三出複葉の本葉が少数出る。この種が属するケシ科キケマン属では、単子葉性への移行が少なくとも2回起こったと推定される。
双子葉・地下子葉性の芽生え
ソラマメ・エンドウ・ベニバナインゲン・アズキ(いずれもマメ科)などでは、子葉は種皮をかぶったまま地中にとどまり(地下子葉性/地下発芽)、貯えている養分を芽生えの他の部分に供給して役割を終える。胚軸はあまり伸びず、子葉の上の茎が伸長して本葉を光の当たるところに持ち上げる。芽生えの光合成は、地上子葉型と違って、最初から本葉が担う。



エンドウ(マメ科)の芽生え。子葉は種皮を被ったまま地中に残り、子葉の上の茎が伸長して地表に出る。
双子葉・中間型の芽生え


インゲンの芽生え。本葉が伸長したころには子葉は黄変して萎びる。
ダイズ(マメ科)・インゲン(マメ科)の芽生えは中間型で、地下子葉性に近いが、地上子葉性の特徴を少しだけ持っている。胚軸が少し伸び、子葉は地上に出て種皮を脱ぐが、種子の中にあったときからあまり展開・成長しない。光が当たって緑色を帯び、短い期間光合成をすることもあるが、早いうちに養分を他の部分に送ってしなび、取れてしまう。

単子葉植物の胚と芽生え(左図)では、子葉は1枚で、子葉の基部が広がって幼芽と幼根をすっぽりと覆う鞘状の構造をつくる。幼芽を覆う部分を「子葉鞘」または「幼葉鞘」[coleoptile]、幼根を覆う部分を根鞘という。幼根は根鞘の先端を突き破り、幼芽は子葉鞘先端近くのすきまから外に出て来る。子葉の先端は胚乳につながり、吸器としてはたらく。
| 1 | 胚軸 |
| 2 | 幼芽 |
| 3 | 幼根 |
| 4 | 子葉鞘 | 子葉 |
| 5 | 根鞘 |
| 6 | 子葉先端部 |
| 7 | 種皮と胚乳 |
単子葉植物の芽生えにも、子葉先端部が地上に出る地上子葉性と地中に留まる地下子葉性とがある。
単子葉・地上子葉性の芽生え
ユリの仲間(ユリ科)・ネギの仲間(ヒガンバナ科)では、子葉先端部が伸長して種皮をかぶったまま地上に突き出す。子葉鞘はあまり伸長せず、幼芽は地表近くで子葉鞘のすきまから出てくる。




タカサゴユリ(ユリ科)の芽生え



タマネギ(ヒガンバナ科)の芽生え
単子葉・地下子葉性の芽生え


ツユクサ(ツユクサ科)の芽生えでは、子葉先端部と子葉鞘の両方が長く伸びる
ツユクサ(ツユクサ科)・キショウブ(アヤメ科)・トウモロコシ(イネ科)とも、種子と子葉は地中に留まって、子葉鞘が伸長して地上に先を出し、幼芽は子葉鞘のすきまからでてくる。ツユクサ・キショウブでは、子葉先端部も伸長するため子葉鞘と種子は離れている。


キショウブ(アヤメ科)の芽生え。1―種皮; 2―子葉先端部; 3―子葉鞘+根鞘; 4―幼芽基部; 5―幼根; 6―不定根
イネ科では、胚は種子の縁にあり、子葉先端部は肥大して胚の他の部分と胚乳の間を充たし、「胚盤[scutellum]」と呼ばれる。胚盤は、ツユクサやキショウブの子葉先端部とは違って伸長せず、種子の中に納まったまま、胚乳から養分を吸収して芽生えの他の部分に供給する。


トウモロコシ(イネ科)の種子の断面。1―内乳 2―胚盤(子葉先端部) 3―幼根(根鞘に包まれている) 4―幼芽(子葉鞘に包まれている)


トウモロコシの芽生え。子葉鞘の上半は緑色になり、下半(根鞘)は先端が破れて幼根が出て来る。続いて、子葉鞘から葉が突き出だし、茎の基部から不定根が子葉鞘を突き破って出て来る。
2-5. 土を突き抜けるしくみは、シュートと根で違う
土壌は、発芽に必要な水分や栄養の供給源だが、生長を始めた芽生えが最初にくぐり抜けなくてはいけない障害物でもある。芽生えの茎頂や根端は、細胞壁が薄くてやわらかい分裂組織を守りながら土を突き抜けるしくみを持っている。同じようなしくみは、多年草が越冬した後で、地下茎や貯蔵根から出てくる茎頂や根端にも見られる。
茎頂
茎頂が土を突き抜けるしくみは「茎頂の屈曲」「鞘状の葉」の2つだ。

ニラ(ヒガンバナ科)の芽生え |
 |
もやしのように、土の中から顔を出すときに、分裂組織の少し下で茎が180度近く曲がって「逆さにした"U"の字」形になるものは多い。ホースか何かで試して見ると実感できるとおり、屈曲点の部分は単純ながら力学的に強い構造が出来る(屈曲点の部分は組織もやや硬くなっている)。茎が伸びると屈曲点の部分が先頭になって土を押しのけて行く。屈曲点が地表からある程度上に出てから、分裂組織がある部分が土から離れ、屈曲が解除されて、最終的には分裂組織が先端に来るようになる。
茎頂が鞘状の葉に守られながら土を押しのけて地中を進み、地表に出た後で鞘のすきまから顔を出すものもある。上で述べたように、単子葉植物の多くでは子葉鞘が同じ役割を担う。

 左: シラン(ラン科)の芽出し。地上に出た濃紫色の鞘状の葉の中から緑色の葉が伸び出す
左: シラン(ラン科)の芽出し。地上に出た濃紫色の鞘状の葉の中から緑色の葉が伸び出す
右: エンゴサク類(ケシ科)の地下部。地中のイモ(球茎)から伸びる白い茎の先端には二つ折りになった白く堅い葉(鞘状の鱗片葉)がついている。鱗片葉が地表に達すると、鱗片葉が開いて、鱗片葉に包まれていた茎や葉、花序が成長する
チューリップのように、地上で展開する葉自体が堅い先端部をもち、鞘と同じ役割を果たすものもある。


チューリップ(ユリ科)の葉は鞘状の堅い先端をもつ
根端
 タマネギ根端の縦断面
タマネギ根端の縦断面
根を作る分裂組織は、茎や葉を作る分裂組織と違って、一生を通じて土と戦い続けることになる。だから、屈曲や鞘のような一時的な方法は使えない。
根端には「根冠」[root cap]という帽子のかたちをした組織がかぶさっていて、分裂組織を守っている。根冠を構成する細胞はやや硬めで、根端が土の中を伸びるにつれて外側から徐々に脱落し、分裂組織から内側に新しい細胞が供給されて補われる。根冠の表面は粘液を出していて(下のダイコンの根端の写真を参照)、それが土の中を伸びるときの潤滑油として働いている。


ダイコン(アブラナ科)芽生えの幼根(左)と根冠(右)
テキスト目次に戻る
ホームに戻る
