
セルロースはβ-グルコース分子が直鎖状に重合した多糖で、強靱で酸や塩基にもかなりの抵抗性があり、水に溶けない一方で高い吸水力をもつ。植物体の乾燥重量の1/3~1/2を占め、天然高分子としては最も大量に存在する。節足動物の外骨格や菌類の細胞壁の主成分であるキチンは、β-グルコース分子とよく似たN-アセチルグルコサミン分子を主な要素とする直鎖重合体だ。
巨大な樹木も、ごく小さな草花も、植物のからだは、たった2種類の単位、
で出来ている。単位の数と配置を変え、そして、ときに個々の単位に特殊な形とはたらきを持たせることで、さまざまな生き方をしている。例えば、花は有性生殖器官として特殊化したシュート、イモはシュートあるいは根が貯蔵器官として特殊化したものだ。
 植物体の模式図。左の図では、個々のシュート(青)・根(赤)を矢印で表わしている。矢印の先端部に頂端分裂組織がある。右の図では、青く塗りつぶされた部分がシュートの集まり(シュート系)、赤く塗りつぶされた部分が根の集まり(根系)。中間の白い部分(胚軸と子葉)は、実質的にはシュートの一部としてはたらくことが多く、シュートの基部と見なされることもある。
植物体の模式図。左の図では、個々のシュート(青)・根(赤)を矢印で表わしている。矢印の先端部に頂端分裂組織がある。右の図では、青く塗りつぶされた部分がシュートの集まり(シュート系)、赤く塗りつぶされた部分が根の集まり(根系)。中間の白い部分(胚軸と子葉)は、実質的にはシュートの一部としてはたらくことが多く、シュートの基部と見なされることもある。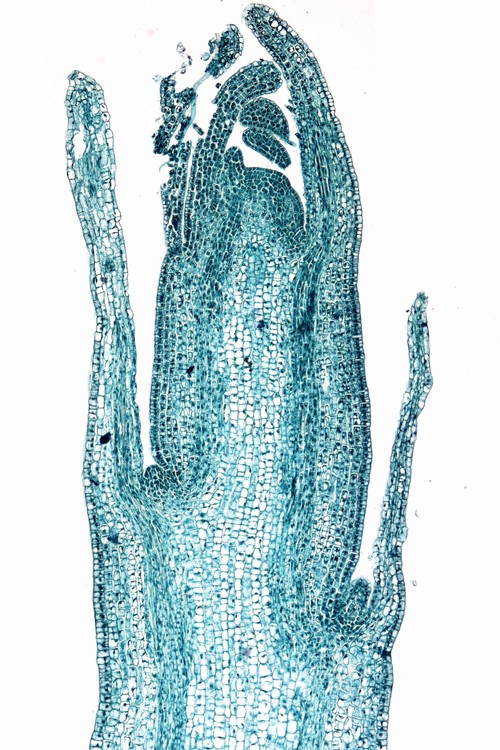
 左―ジャガイモ(ナス科)の茎頂; 右―タマネギ(ユリ科)の根端の縦断面。分裂組織の細胞は(ファストグリーンでよく染まっている)は、液胞の量が少なく、細胞に占める細胞質ゾルの割合が高いため濃く染色される。
左―ジャガイモ(ナス科)の茎頂; 右―タマネギ(ユリ科)の根端の縦断面。分裂組織の細胞は(ファストグリーンでよく染まっている)は、液胞の量が少なく、細胞に占める細胞質ゾルの割合が高いため濃く染色される。
シュート・根はどちらも「極性をもつ軸構造」──つまり、2つのはじっこ、先端(頂端)[apex]と基部[base]がある「棒」だ。頂端には、分裂組織[meristem](細胞分裂が活発な部分)があり、頂端分裂組織[apical meristem]という。シュートの頂端分裂組織を茎頂分裂組織(シュート頂分裂組織)[SAP; shoot apical meristem]、根の頂端分裂組織を根端分裂組織[RAP; root apical meristem]という。
シュート・根は、頂端分裂組織で出来た細胞が付け加わることで伸長する。頂端分裂組織から離れるほど、古い細胞で構成されているから、シュートや根を空間的に先端→基部へとたどることは、時間的に現在→過去へとたどることにもなる。頂端から離れるにつれて分裂の頻度が落ちて、代わりに、一つ一つの細胞が軸方向へ伸長する。さらに頂端から離れると、細胞の伸長も余りしなくなり、細胞や組織は、働きに応じた形態の違い(分化[differentiation])がはっきりとしてくる。

茎頂分裂組織は、茎を伸ばすと同時に、葉を作り出すはたらきもする。だから、1本のシュートの茎と葉は、同じ頂端分裂組織によって作られる。このことは、茎と葉をまとめてシュートという単位と見なす大きな理由の1つだ。
 分裂組織の主要な3タイプ。左―頂端分裂組織・中―介在分裂組織・右―側方分裂組織(形成層)
分裂組織の主要な3タイプ。左―頂端分裂組織・中―介在分裂組織・右―側方分裂組織(形成層)1つの植物が持つシュートをまとめてシュート系[shoot system]、根をまとめて根系[root system]と呼ぶ。つまり、1つの植物は、シュート系と根系に大別される。
2種類の単位が積み上がることで身体ができる植物は、身近な動物(多細胞動物)と、次のような点で違っている。
植物の成長は、単位の継ぎ足し+個々の単位の成長だ。

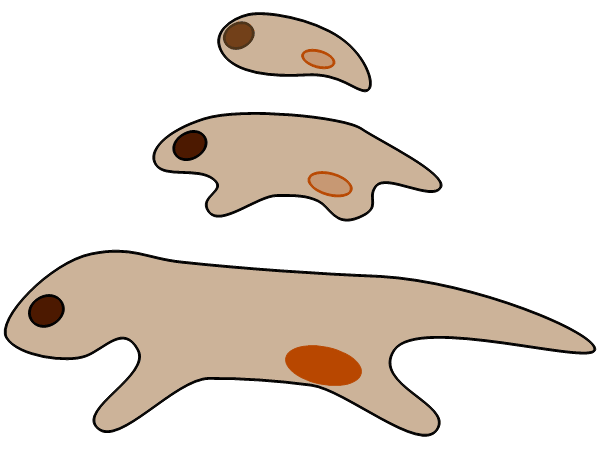
脊椎動物や節足動物をはじめ、身近な動物の多くは、発生の初期に器官の分化を終える。そのため、(種類が同じであれば)1個体当たりの器官の数(眼の数、足の数、など)が決まっている。植物では、成長につれて新しいシュートや根が、次々と継ぎ足されて(付け加わって)いくので、大形個体の方が多数の単位からなることが多い。
成長様式の違いは、「サイズに関わらず同じ食物(光と土壌養分)」「多くの食物を取るのには、動くよりも占有面積を広くする方が有利」という植物のくらし方を反映していると考えられる。また、動物のように成長に応じて食物を変える必要に迫られることがないことは、変態[metamorphosis](成長過程での劇的なモデルチェンジ)が、植物ではほとんど見られない理由の1つだろう。
植物のからだのサイズの個体差が、動物に比べて大きい(このことを「サイズの可塑性が大きい」と表現することもある)ことも、成長様式の違いによって説明できる。
 セイヨウカラシナ(アブラナ科)のすぐ近くに生えていた大小2個体。大きい方の個体は、写真に収まりが良いように三つ折りにしてある。中央右に小さい方の個体がある。大きい方の個体は、シュート・葉・茎の数が多く、1つ1つのサイズも大きい。茎の長さで見ると両者の比は約5倍、花の数で見ると少なくとも100倍はある。
セイヨウカラシナ(アブラナ科)のすぐ近くに生えていた大小2個体。大きい方の個体は、写真に収まりが良いように三つ折りにしてある。中央右に小さい方の個体がある。大きい方の個体は、シュート・葉・茎の数が多く、1つ1つのサイズも大きい。茎の長さで見ると両者の比は約5倍、花の数で見ると少なくとも100倍はある。動物と植物では、有性生殖器官の作られ方・使われ方に違いがある。
| 動物 | 植物 | |
|---|---|---|
| 有性生殖器官 | 卵巣・精巣・交尾器など | 花/果実・花序/果序 |
| 数 | 雄1セット、雌の1セット あるいは雌雄1セットずつ |
1~複数 多くの場合、個体が大きいほど多くなる |
| 作られ方 | 胚の段階で1回作られる | 成長過程で次々と枝先に作られる |
| 使われ方 | 同じ生殖器官が一生の間「再利用」される | 1回使われた生殖器官は「使い捨て」られる 次の繁殖期には新しい生殖器官が使われる |
このことは、植物の性別(雌雄性)を、動物と比べものにならないほど複雑で多様なものにしている。
動物では、器官の細胞は分化を終えていて、他の器官の細胞を生み出す能力が乏しくなっている。植物のシュートや根の細胞は、植物体のあらゆる器官の細胞を生み出す能力(分化全能性)を失っておらず、一部のシュートや根を身体から切り離し、切り離された単位が新しい植物に再生することで無性生殖(クローン繁殖)を行うものが多い。農民や園芸家は古くからこの性質を利用してきた(挿し木・挿し芽・茎頂培養・多くのイモ類の増殖法)。
休眠(活動に適さない時期を、一時的活動停止状態で過ごす)のときには、活動に適した部分を切り離して休眠に適した部分だけを残す。樹木の落葉や、草がイモや球根を残して枯れるのは、この例。
動物における神経系・循環系・消化器系など、別々の器官どうしを横断的につなぐしくみは、維管束のつながりが大部分を担っている。動物のように筋肉や神経系によって全身を瞬時に統合して働く仕組みはつくれないが、維管束を通じた情報伝達には、小形の植物体なら数分で全体に及ぶものもあることが知られている。
| シュート系 | 根系 | 葉緑素 | はたらき | |
|---|---|---|---|---|
| 地上部 | 多数 | 気根 | + |
|
| 地下部 | 地下茎 | 多数 | - |
|
植物体は、生長単位によってシュート系[shoot system]と根系[root system]に大別できる。また、生活上の役割分担によって地上部[aerial part]と地下部[underground part]に分けることができる。「シュート系=地上部」で「根系=地下部」だったら話は単純で分かりやすく、実際にそういう植物もあるが、そうでない例も無数にある。
 ジャガイモ(ナス科)の地上部と地下部。中心軸は地上・地下ともシュートで、所々から貯蔵用のシュート(イモ)と根を出している。
ジャガイモ(ナス科)の地上部と地下部。中心軸は地上・地下ともシュートで、所々から貯蔵用のシュート(イモ)と根を出している。
多年草の多くは地下茎[underground stem]を持っていて、養分を蓄える役割をしている。地下茎は棒状のときもあるけれど、丸いイモのようになっていたり(ジャガイモなど)、ややこしい形の塊になっている(ショウガなど)ときもある。
地上部の根を「気根」[aerial root]という。地下茎ほどふつうではないが、植物体を支える(支柱根)、何かに付着する(付着根)など、さまざまな例がある。



シュート系と根系、地上部と地下部の比率は、次のような制約を受ける。
だから、やたらと片方ばかり多くなることはふつう出来ないし、植物の種類や生育している環境によっても変わる。例えば、海岸砂丘のような土壌が不安定で水分が少ないところに生えている植物は、多数の長い根をつけて水分と力のバランスを保つことが多い。また、同じ植物でも、乾燥状態で育てると根系の比率が高くなり、逆に湿潤状態で育てるとシュート系の比率が高くなる。鉢植えしている植物では、二、三日水やりをさぼると、盛んに新しい根を出し始めることがある。
草木の植え替えのときにはどうしても根系に損傷を与えてしまうので、葉が落葉したり、シュートが部分的に枯れて、バランスを取ろうとする。しかし、悪くすると植物全体が枯れてしますことがあるので、シュート系が休眠している時期に行ったり、シュート系を刈り込んだり、多めに水やりをすることが多い。挿し木のときに、葉を切り取ってやるのも同じ理屈だ。
植物の器官(例えば、葉・茎・根・花びら・雄しべ・種子)を薄切り(切片 せっぺん [section])にして顕微鏡で観察すると、たくさんの細胞の集まり(組織)が見える。こういうときの観察のポイントは、大きく分けて3つある。
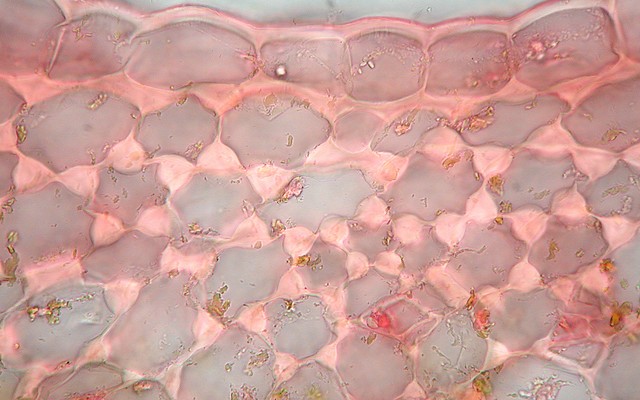
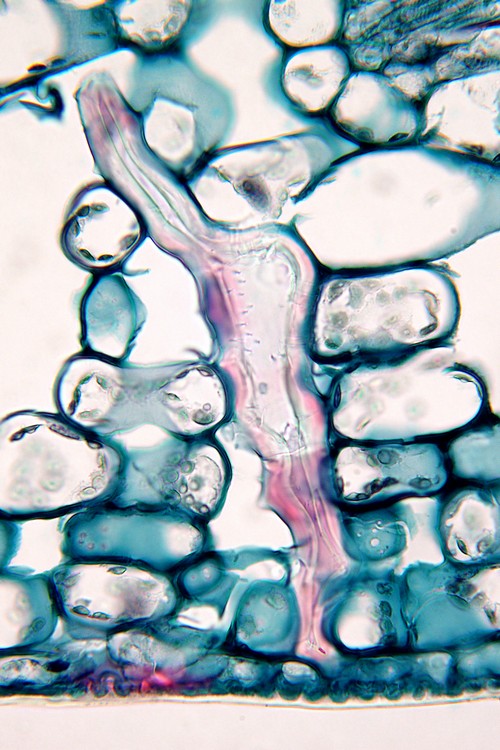 ヤブツバキ(ツバキ科)葉の海綿状組織中に見られる異形細胞。縦長の不規則形で厚い細胞壁をもつ。
ヤブツバキ(ツバキ科)葉の海綿状組織中に見られる異形細胞。縦長の不規則形で厚い細胞壁をもつ。
植物の組織の多くは、形状や機能が似通った細胞の集まりだ。しかし、他の細胞と形状が異なる細胞が混じっている場合がある。そのような細胞を異形細胞[idioblast]という。
植物細胞は、原則として、細胞膜で囲まれた細胞質[cytoplasm]の外側に細胞壁[cell wall]を作る。細胞壁はセルロース・ヘミセルロース・ペクチンを主成分とする細胞外マトリックスで、弾性と伸縮性がある。
セルロースはβ-グルコース分子が直鎖状に重合した多糖で、強靱で酸や塩基にもかなりの抵抗性があり、水に溶けない一方で高い吸水力をもつ。植物体の乾燥重量の1/3~1/2を占め、天然高分子としては最も大量に存在する。節足動物の外骨格や菌類の細胞壁の主成分であるキチンは、β-グルコース分子とよく似たN-アセチルグルコサミン分子を主な要素とする直鎖重合体だ。
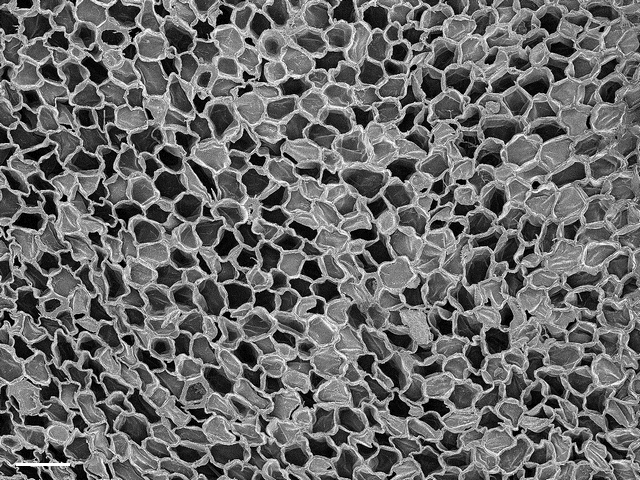
 コルクの断面に見られる細胞壁(SEM像・スケールは50μm)
コルクの断面に見られる細胞壁(SEM像・スケールは50μm)
細胞壁は、隣接する細胞の細胞壁と密着しているか、または、空気で充たされたすきま(細胞間隙)と接している。隣接する細胞どうしの細胞質は、細胞壁を貫通する経路である原形質連絡でつながっている。
 ジャガイモ(ナス科)のごく初期の芽(中央のドーム)付近の断面。ファストグリーン(染色液)によって細胞質は濃緑色に染色されるが、液胞は染色されない。細胞分裂と成長がさかんな芽では細胞全体が濃く染色され、周辺部の染色度の低い細胞ははっきり違う。
ジャガイモ(ナス科)のごく初期の芽(中央のドーム)付近の断面。ファストグリーン(染色液)によって細胞質は濃緑色に染色されるが、液胞は染色されない。細胞分裂と成長がさかんな芽では細胞全体が濃く染色され、周辺部の染色度の低い細胞ははっきり違う。
活発に細胞分裂や細胞成長をしている組織では、細胞壁は薄くてあまり強度に貢献しない。また、液胞の占める容積は少なく、細胞の大部分はサイトゾル(細胞質基質)で充たされている。
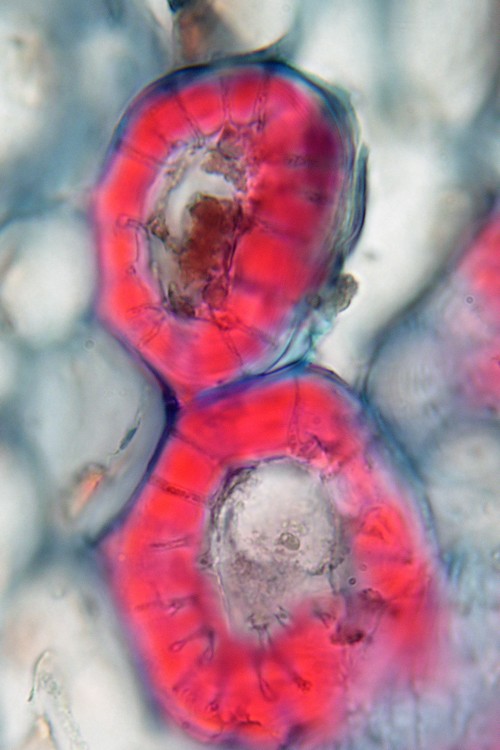
 右: リュウキュウアサガオ(ヒルガオ科)の花冠表皮。花色色素のアントシアニンを貯える紫色の液胞が細胞の大部分を占める。
右: リュウキュウアサガオ(ヒルガオ科)の花冠表皮。花色色素のアントシアニンを貯える紫色の液胞が細胞の大部分を占める。
成長を終えた細胞では、細胞壁がやや厚くなり、液胞が細胞の容積の多くを占めて、サイトゾルは細胞膜に沿って分布する。液胞は、さまざまな水溶性物質の「保管庫」であるとともに、サイトゾルの体積に対して細胞の体積や表面積を大きく保つ(空間充填機能)。水分条件によって細胞の体積や濃度は変化するが、変化の多くは液胞が負い、サイトゾルの体積・濃度の変化は緩和される。
水が十分にある条件下では、膨らもうとする細胞を細胞壁が押さえつけて、空気とタイヤ、あるいはチューブとタイヤのような緊張関係を保ち、細胞の生長の可能性を残しつつ細胞に強さを与えている。水が不足すると液胞が収縮→細胞が収縮して、すぐに萎れてしまう("水涸れ")が、水不足が解消されれば元に戻る。
 植物細胞のモデル。細胞壁・サイトゾル(細胞質基質)・液胞だけを示す。水が十分に供給されているとき(左)は、細胞が膨らむ力(1)と細胞壁の反発力(2)が釣り合っている。水が不足すると(右)、液胞が収縮して細胞と細胞壁の押し合いがなくなり、組織の強度が下がる。
植物細胞のモデル。細胞壁・サイトゾル(細胞質基質)・液胞だけを示す。水が十分に供給されているとき(左)は、細胞が膨らむ力(1)と細胞壁の反発力(2)が釣り合っている。水が不足すると(右)、液胞が収縮して細胞と細胞壁の押し合いがなくなり、組織の強度が下がる。

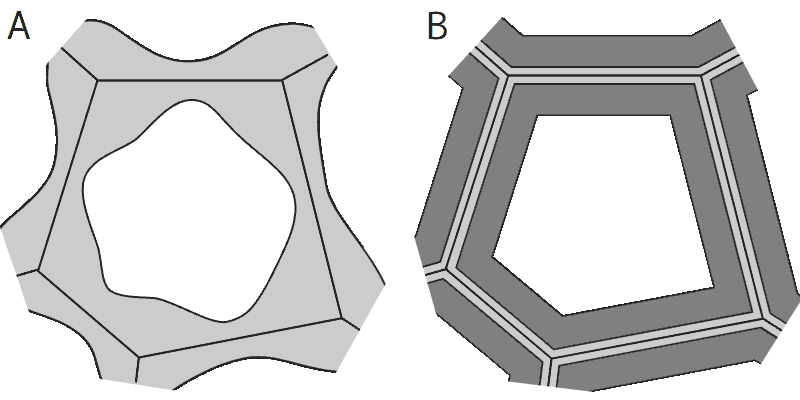
さらに強度が必要な細胞では、もともとの細胞壁(一次細胞壁―左図では淡灰色の線で表示)の内側に追加の細胞壁(二次細胞壁―左図では濃灰色で表示)が服の裏地のように付け加わることでもっと厚くなるが、その分生長の余地は少なくなる。
厚角細胞(A)と厚壁細胞(B)の模式図
二次細胞壁の成分組成が一次細胞壁とあまり変わらないような細胞を厚角細胞(こうかくさいぼう)と呼ぶ(細胞の角のところが特に厚くなることが多いため)。厚角細胞は、ある程度の細胞の成長も可能で、細胞も生きていることが多い。

細胞生長の必要性が少なく、柔軟性よりも剛性が必要な組織(硬くなったとげ・木の材・熟した種子や果実の皮など)には、厚壁細胞(こうへきさいぼう)と呼ばれる細胞が見られる。厚壁細胞の二次細胞壁はリグニンを多く含み、伸縮性を失っており、細胞自体は死んでいることが多い。厚壁細胞は形状によってさらに細かく分類される。例えば、

 ボケ(バラ科)の果実断面・果皮付近。石細胞が散在する。
ボケ(バラ科)の果実断面・果皮付近。石細胞が散在する。
厚角細胞・厚壁細胞は、ボケ果肉の石細胞やヤブツバキ葉の不規則形の細胞のように異形細胞として存在する場合もあるが、集合して「~組織」と呼ばれる場合の方が多い。厚角細胞から構成される組織は厚角組織[collenchyma]、厚壁細胞からなる組織は厚壁組織[sclerenchyma]と呼ばれる。細胞壁が薄いままの細胞(柔細胞)からなる組織は、柔組織[parenchyma]と呼ぶ。




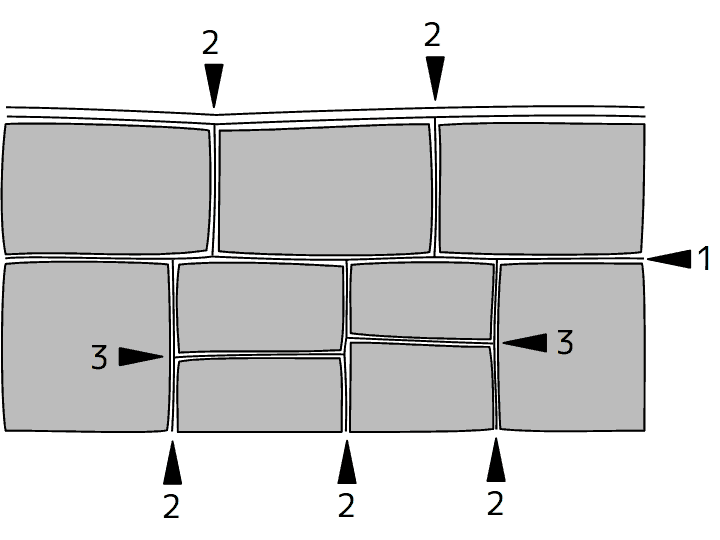 植物細胞の配列の模式図。矢頭と数字は、細胞分裂の方向と相対的な順序を示す。
植物細胞の配列の模式図。矢頭と数字は、細胞分裂の方向と相対的な順序を示す。
隣り合った細胞は、両方の細胞からつくられた2枚の細胞壁でつながっている。細胞分裂のときには、出来た2つの細胞は新しくできた細胞壁で互いにつながり合うことになる。だから、植物組織での細胞の並び方は細胞分裂[cell division]の順序と方向で決まり、逆に言うと、細胞の並び方を見れば、分裂の順序と方向をある程度推定することが出来る。
 タマネギ(ユリ科|ヒガンバナ科)の根の縦断面に観られる細胞列。
タマネギ(ユリ科|ヒガンバナ科)の根の縦断面に観られる細胞列。
細胞分裂が常に同じ方向でだけ起こると、細胞は一列に並ぶことになる。若い根の組織が好例。

細胞分裂がある一平面上の細胞数を増やすような方向で繰り返されると、細胞がその面に整然と並ぶことになる。断面を見ると、このような分裂を垂層分裂[anticlinal division]と言う。多くの植物器官では、表皮細胞は垂層分裂を繰り返すので、表皮はその内側からはっきりと区別された細胞層になる。細胞がしっかりと密着して外と中の仕切りになるためには、このような分裂パターンの方が適しているから、このことは理に適っている。組織によっては複数の細胞層が積み重なった構造をとることも多い。垂層分裂と直交する方向で起こる細胞分裂を並層分裂[periclinal division]、どちらとも言えないような分裂を斜分裂[oblique division]という。また、縦長の細胞で構成された細胞層を柵状組織[palisade; palisade parenchyma]と呼ぶ。
細胞分裂がさまざまな方向で行われると、出来た組織に細胞の配置も不規則なものになる。断面では、細胞はまるで野面積みの石垣のような並び方をしている。このような組織では、細胞分裂後の細胞の成長により、細胞間にすきま(細胞間隙 [intercellular space])が出来ることが多い。すきまが特に多い、すかすかな組織のことを海綿状組織[spongy tissue; spongy parenchyma]と呼ぶ。多くの植物器官ではこのような、方向性のない組織がもっとも多くの体積を占めている。
胚嚢や種子の内乳(胚乳の一種)では、細胞核だけが分裂を繰り返し、核がたくさんに増えて後で細胞膜や細胞壁が作られる(植物の種類によってはそうでないのもある)。こういう場合には、細胞の配列からは細胞分裂の順序や方向は分からない。胚嚢では、大きな細胞の中に小さな細胞が何個か(ふつうは6個)埋め込まれている。花粉も、大きな細胞の中に1個か2個の小さな細胞が埋め込まれている。
植物が緑に見えるのは細胞内の葉緑体による。花びらや果実のように緑色以外の色をしている器官や、地下器官のように光が当たらない器官は、葉緑体のない細胞からなっている。


緑色に見える器官でも、葉緑体は器官の表面に近い、光が届くところに局在し、茎や雌しべのようなある程度厚みがあるものでは、中の方は葉緑体がない細胞がほとんどだ。


葉のように薄い器官では、表皮と維管束以外の細胞はほとんどが葉緑体をもっているが、キャベツや白菜のように幾重にも重なっているときには内側の方へ行くほど緑色が薄まり、最後には白っぽくなる。また、多肉植物の乾燥に適応した厚い葉では、内部の組織は葉緑体をもたない。
上で「葉緑体がない」というのは間違ってはいないが不十分な記述だ。分裂組織の細胞は葉緑体に分化可能な細胞小器官(原色素体[proplastid])をもっており、細胞分化の過程で、葉緑体・デンプンを貯蔵するアミロプラスト[amyloplast]・カロチノイドを貯蔵する有色体/クロモプラスト[chromoplast]など、多種多様な機能と形状をもつ細胞小器官に分化する。原色素体と原色素体から分化した細胞小器官を総称して色素体/プラスチド[plastid]という。葉緑体がない植物細胞も、何らかの色素体をもつことが殆どだ(多くの被子植物の精細胞のように、色素体をもたない細胞もある)。
キャベツや白菜では、外側の葉が生長して反り返ると、光が当たるようになった内側の葉では、原色素体で葉緑素が合成されて葉緑体となり、緑色の葉に変わっていく。明るいところに置いてあったジャガイモがうっすらと緑色になるのも、同じ理由による。枯れた葉では、葉緑体の葉緑素が分解されていくため、やはり緑色は失われていく。
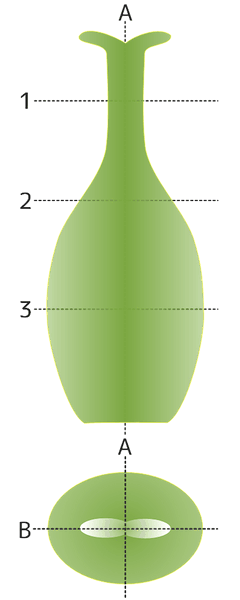 雌しべの横断(1~3)と縦断(A・B)
雌しべの横断(1~3)と縦断(A・B)
単細胞生物や、単離された細胞は、そのまま観察することが出来るが、組織の中の細胞を観察するためには、組織を薄切り(切片)にして、光が十分透過するようにしなくてはならない(場合によっては、平たく押しつぶして観察する場合もある)。切片は、高倍率で観察するほど薄くする必要があり、光学顕微鏡の切片は1~100μm(マイクロメートル)、透過型電子顕微鏡では0.1μm=100nm(ナノメートル)以下の厚さに切る。
薄切りにする方向は、目的に応じて決める。軸に対して垂直に切ることで得られる切片を横断切片、軸に平行に切って得られる切片を縦断切片という。
生物の組織は切片にすると透明なものが多い。ただ、切片が厚いときは、物質によって光の透過率や屈折率が違うので、だいたいは問題なく観察できる。
光学顕微鏡の場合、薄い切片では色素によって染色することが必要なことが多い。色素によって染まりやすい物質が違うので、複数の色素を使えば、見やすくなるだけでなく、物質の違いが色で分かるようになる。例えば、植物では、細胞核を染めるヘマトキシリン、リグニンやセルロース(とりわけリグニン)を染めるサフラニン、細胞質やセルロースを全体的に染めるファストグリーンを組み合わせた三重染色がよく使われる。
色素の染まり易さは、物質との化学的な相性によって決まることが多いが、抗原抗体反応(タンパクと免疫グロブリンの反応)や相補的な核酸一本鎖どうしの結合(インサイテューハイブリダイゼイション[in situ hybridization]; 略して「ISH」いっしゅ)を使うと、ある特定のタンパクやDNA・RNAだけを染めることができる。この方法は、分子レベルでの生物学研究では大変有効な方法で、盛んに用いられている(特に、下の蛍光顕微鏡と組み合わせることが多い)。
色素として蛍光色素(光が当たると、当たった光と違う色の光(蛍光)を発する特性がある物質。蛍光色素を含んだ塗料が蛍光塗料)、最初に当てる光(投射光)にはエネルギーが大きい紫外線を使い、フィルターで蛍光だけがみえるようにすると、プレパラートの中で蛍光色素があるところだけが輝いて見える。紫外線を投射する装置とフィルターを備えた顕微鏡を「蛍光顕微鏡」という。
透過電子顕微鏡の場合、切片が非常に薄いので、色素染色はほとんど意味が無く、屈折率の差も非常に小さい。ウランや鉛のような原子量の大きい金属原子(電子線を吸収する)を組織と結びつけて染色する。