0-3. 陸上植物を構成するグループ
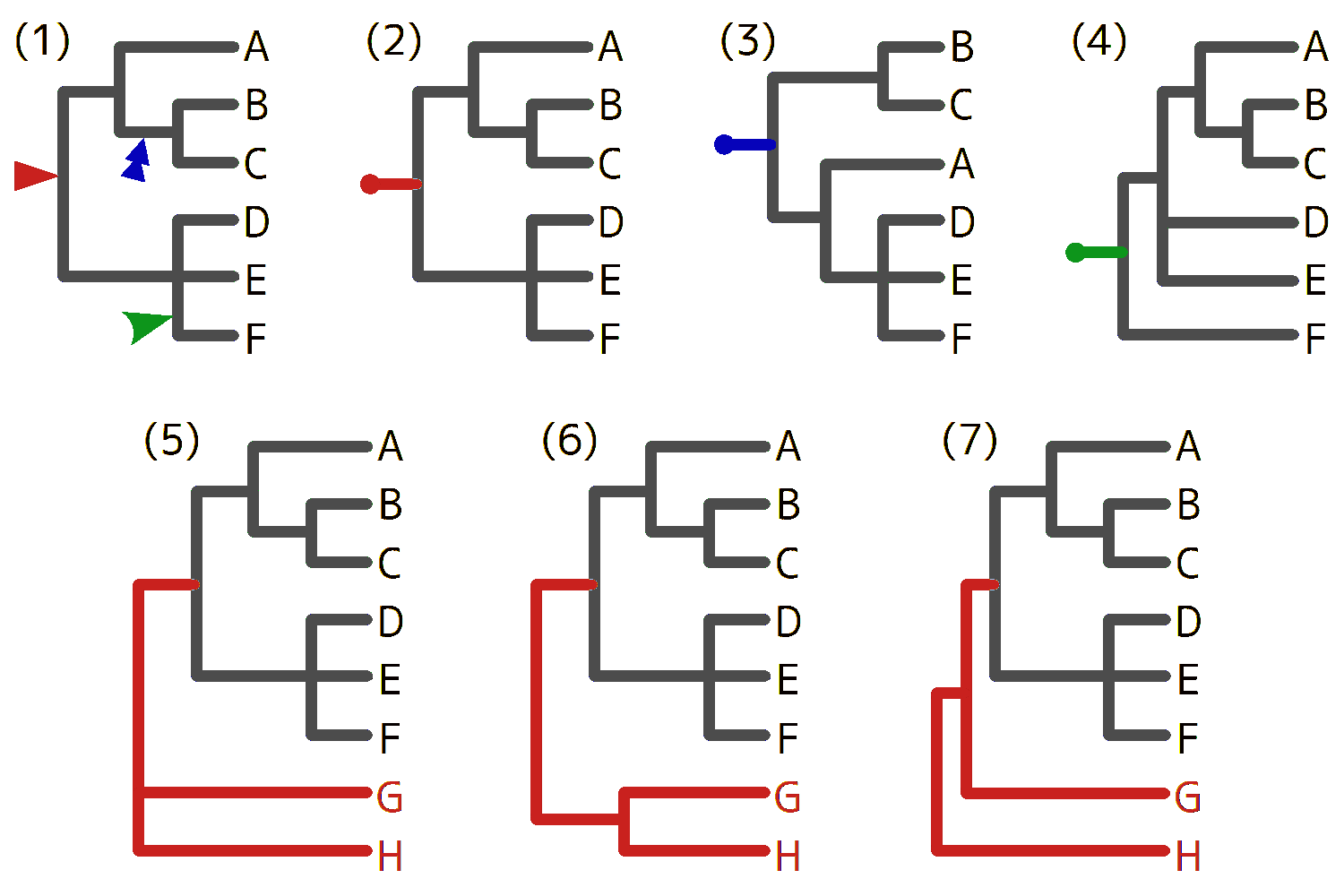
0-3-1. 現生陸上植物のグループと系統関係
現生の陸上植物は、維管束をもたないコケ植物と維管束植物とに分けられ、コケ植物はセン類(蘚類)・タイ類(苔類)・ツノゴケ類の3つのグループに細分される。維管束植物は種子をつくらないシダ植物と種子植物とに分けられ、シダ植物は小葉植物と大葉シダ植物、種子植物は裸子植物と被子植物と、それぞれ2つのグループに細分される。
コケ植物 タイ類 維管束がない 散布体は胞子 配偶体が生活の本体
セン類
ツノゴケ類
維管束植物 シダ植物 小葉植物 維管束がある 配偶体・胞子体とも独立生活
大葉シダ植物
種子植物 裸子植物 散布体は種子 胞子体が生活の主体
被子植物
コケ植物の3群: 左―ジャゴケ(タイ類)・右上―ギンゴケ(セン類)・右下―ナガサキツノゴケ(ツノゴケ類) シダ植物の2群: 左―マンネンスギ(小葉植物)・右―クサソテツ(大葉シダ植物) 種子植物の2群: 左―アカマツ(裸子植物)・右―シロツメクサ(被子植物)
陸上植物では、
配偶体[gametophyte]: 胞子から体細胞分裂ででき、細胞ごとにゲノム1組がある("n"と表現される)。造卵器[archegonium; 複数形 archegonia)]*と造精器[antheridium; 複数形 antheridia]をつけ、造卵器内に♀配偶子(卵細胞)、造精器内に♂配偶子(精細胞)ができる。
胞子体[sporophyte]: 卵細胞と精細胞が融合した受精卵から体細胞分裂ででき、細胞ごとにゲノム2組がある("2n"と表現される)。胞子嚢[sporangium; 複数形 sporangia]をつけ、胞子嚢内に胞子ができる。
陸上植物の卵細胞は造卵器内に留まって受精し、受精卵から初期の多細胞体=胚[embryo]となる。陸上植物の分類体系上の名称「有胚植物」[Embryophyta]はこの特徴に基づいている。被子植物では、卵細胞と2個の助細胞からなる卵装置[egg apparatus]が造卵器に当たるとされている。
の2タイプの多細胞体があり、生活環(個体発生と有性生殖の過程)において
コケ植物(左)・シダ植物(中)・種子植物(右)における配偶体(暗灰色)と胞子体(明灰色)の模式図
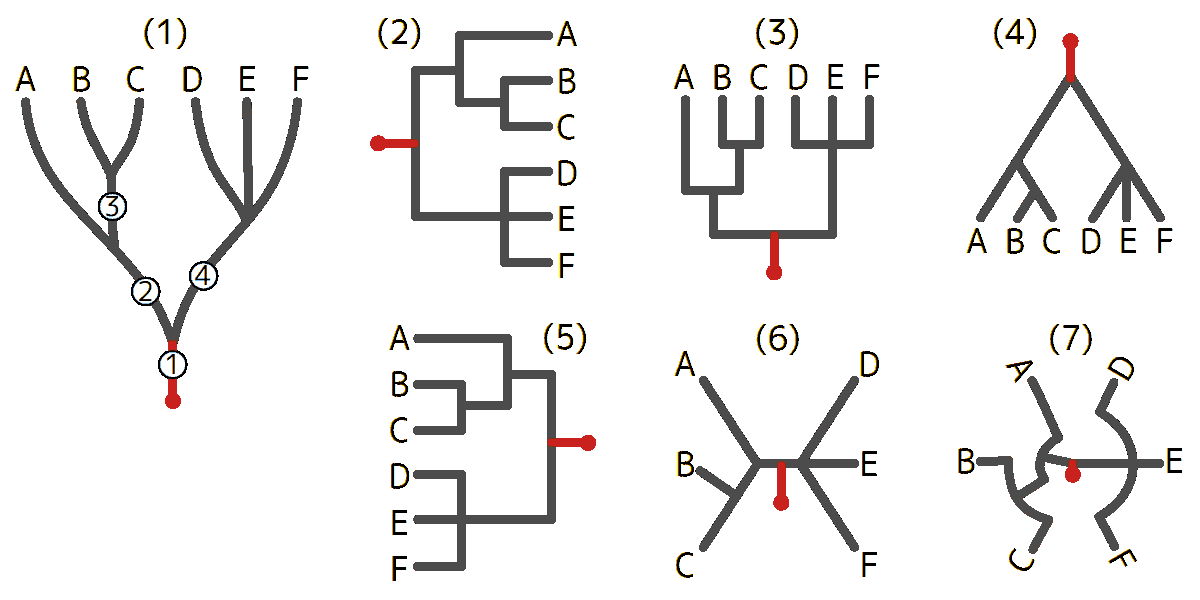
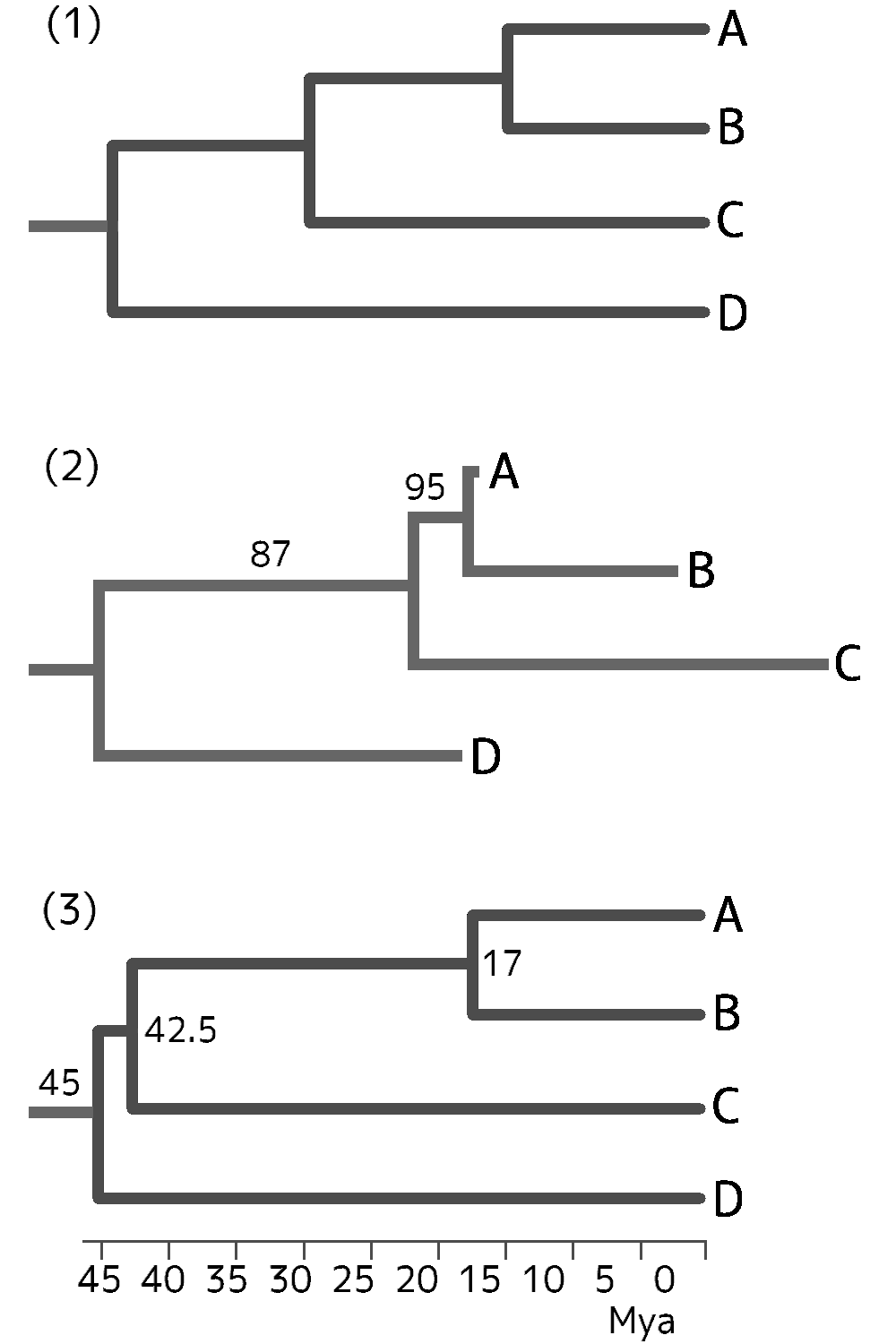
グループ間の系統関係(分岐関係)は、遺伝子の塩基配列に基づいて推定されている。最有力視される仮説を下の系統樹で示す。左端が共通祖先、右端に各グループが並ぶ。
現生陸上植物の系統関係
タイ類 蒴柄植物 コケ植物 陸上植物 小葉植物 ┓ 大葉シダ植物 維管束植物 裸子植物 真葉植物 種子植物
コケ植物・維管束植物・種子植物・蒴柄植物(タイ類+セン類)・真葉植物(大葉シダ植物+種子植物)は、それぞれの共通祖先にあたる枝(図に書き込んだ)に由来する全ての枝(末端も含む)の集まりだ。このような集まりを単系統群[monophyletic group](クレード[clade])という。これらに対して、シダ植物は、単系統群ではなく、2つの単系統群(小葉植物と大葉シダ植物)の複合群と見ることもできるし、単系統群(維管束植物)から単系統群(種子植物)を除いた残りと見ることもできる。単系統群から1個または複数の単系統群を除いたグループを側系統群[paraphyletic group](グレード[glade])といい、シダ植物は典型例の1つだ。
0-3-2. コケ植物と維管束植物
遺伝子の塩基配列から推定された系統樹では、現生植物の祖先系統は、2つの単系統群――配偶体を生活の主体とするコケ植物と胞子体を生活の主体とする維管束植物とに分岐する。
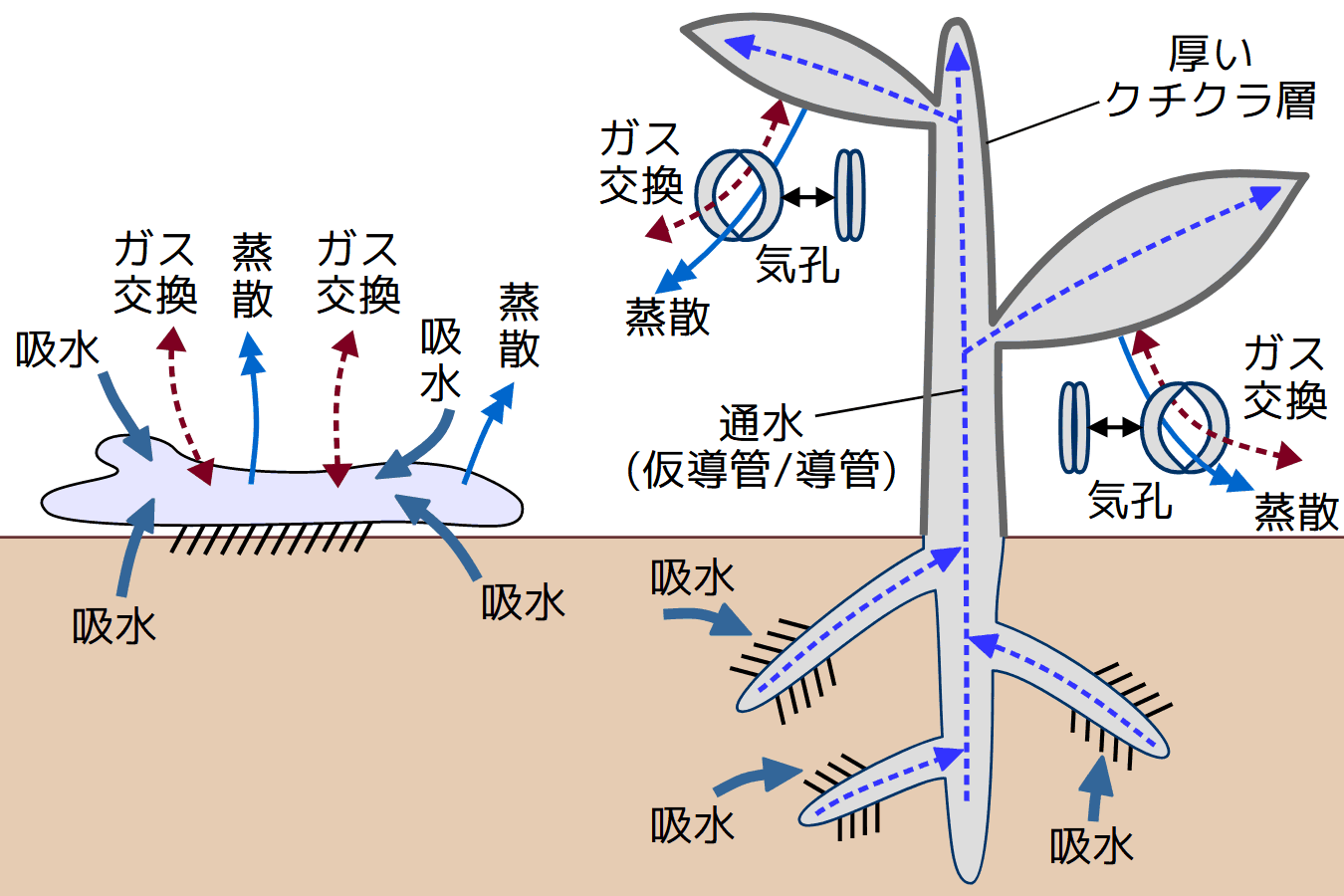
コケ植物配偶体と維管束植物胞子体との最も際立った違いは、光合成を中心とする生命活動に欠かせない吸水とガス交換のあり方だ(下表)。
コケ植物配偶体と維管束植物胞子体との比較
コケ植物配偶体 維管束植物胞子体 模式図 基質への固着 仮根 根 吸水 体表全体 蒸発/蒸散 気孔; 厚いクチクラ層が 通水 なし/リグニンを欠く通水細胞群 維管束; 導管・仮導管の細胞壁は 含水率恒常性 低い(変水性) 高い(恒水性) 乾燥耐性 高い 一部を除き低い 生育型 地表・岩表・樹幹などを 立体的かつ高い植物体が可能
コケ植物では吸水とガス交換は体表全体で起こる。密生する仮根は、毛細管現象で基質から水を吸い上げ、体表からの吸水を助ける。葉状体・茎葉体とも少数の細胞層からなる断面構造をもつため、体表からの水や空気は各細胞に行き渡る。
体表から吸水できるコケ植物配偶体は、周囲が乾燥すると、逆に体表から蒸発によって水分が奪われて乾燥し、生命活動が困難となる。コケ植物の植物体は地表や岩の表面、木の幹などを低く、たいていは平面的に覆うようなかたちとなる。コケ植物の多くには乾燥耐性[Desiccation tolerance; DT]があり、干からびて生命活動を停止しても、雨や霧が掛かると速やかに水分を吸収して活動を再開する。この2点で、コケ植物は、地衣 [lichen](菌類と単細胞緑藻またはシアノバクテリアとの共生体)や、シアノバクテリアの群体が形成するマットと似ている。
乾燥時のタチヒダゴケ(セン類)と霧吹きで湿らせて30分後のようす チヂミバコブゴケ(セン類)の乾燥時(右上)と水漬時(右下) 樹幹のウメノキゴケ類(地衣)。右の写真は、湿潤状態(左)と乾燥状態(右)の色の違いを示す。 地表生のシアノバクテリア、イシクラゲの群体。いずれも、左が乾燥状態で「かさぶた」状、右は湿潤状態でゼリー状。
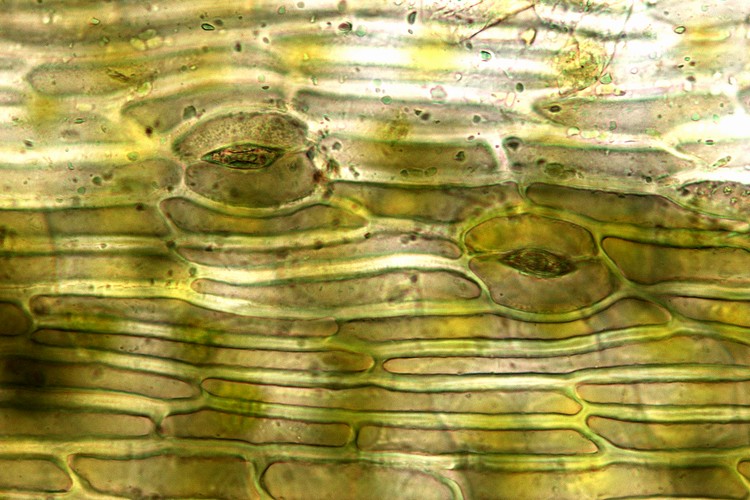
茎葉体を持つコケ植物では、茎の中心や葉の中肋に細長い細胞があり、単純な通水機能を持つ。特に、セン類の一部(内水性セン類 [Endohydric mosses])の茎はクチクラで覆われ、発達した通水細胞群が中心部を貫通しており、水は主に茎の芯を通じて植物体に行きわたる。
維管束植物では、吸水とガス交換の場が分離している。
透水性・通気性が低いクチクラに覆われた体表では、ガス交換と蒸散(水の蒸発)の多くは気孔で行なわれる。気孔は外界の様々な条件に応じて開閉し、ガス交換・蒸散のタイミング・量が調節される。各細胞は細胞間隙を通じて気孔とつながっている。
根は土壌中に伸び、広範囲から水分と養分を集める。また、根は特定の菌類の生育の場となる(菌根)ことによって養分の吸収力を高めている。根で吸水された水(水溶性養分を含む水溶液)は、管状の細胞が連続した通水経路=維管束(仮道管・導管・篩管)を通って茎、さらに葉へと移動する。仮導管・導管の細胞壁は、リグニンを含む二次細胞壁によって補強され、植物体を支持する骨組みとしても機能する。蒸散は、導管・仮導管を水が上昇する原動力となる。
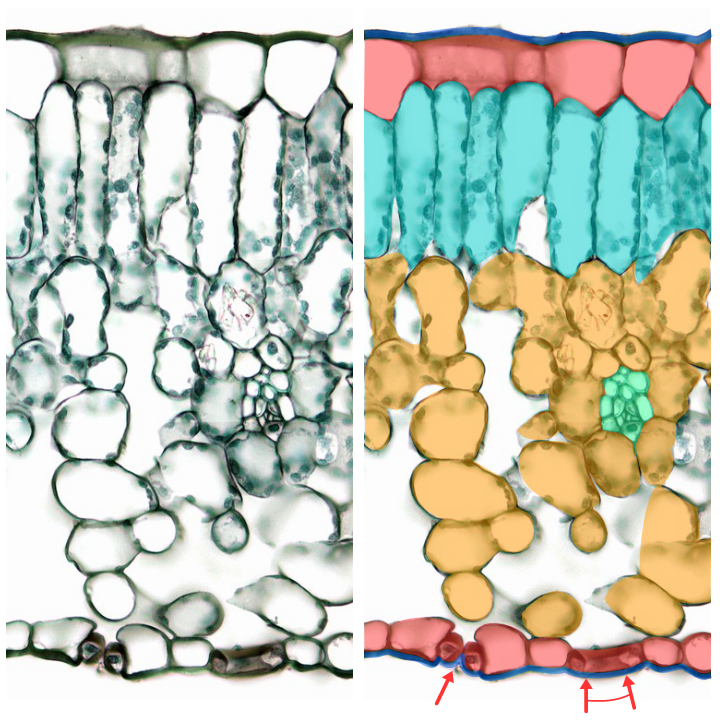
アオカズラ(アワブキ科)の葉の断面(原図と組織ごとに塗り分けた図)
厚いクチクラ・気孔・根・維管束による吸水・ガス交換のシステムによって、維管束植物では、器官レベルではコケよりずっと立体的な細胞の配置、個体レベルではコケよりはるかに大形の、空中に高くそびえ立つ植物体が可能となった。
厚いクチクラ・気孔・根・維管束の進化的起源
クチクラはコケ植物の配偶体・胞子体に広く見られるが、水の蒸発を防ぐほど厚くないことが多い。セン類・ツノゴケの胞子体は比較的発達したクチクラと気孔を持ち、機能ははっきりと分かっていないが、維管束植物のクチクラ・気孔と共通の起源を持つと推定され(Ligrone & al. 2012)、気孔の発生過程・制御遺伝子は、セン類・ツノゴケ・維管束植物を通じて共通性が高い(Qu & al. 2017)。
Ligrone R, Duckett JG, Renzaglia KS. 2012. Major transitions in the evolution of early land plants: a bryological perspective. Ann Bot 109 (5): 851-871. DOI: https://doi.org/10.1093/aob/mcs017 (link )
Qu X, Peterson KM, Torii KU. 2017. Stomatal development in time: the past and the future. Current Opinion in Genetics & Development 2017, 45:1–9. DOI: http://dx.doi.org/10.1016/j.gde.2017.02.001
恒水性・変水性と「乾燥への強さ」
植物体内の水分条件は、外界の水分条件の変動と連動する。連動の度合いが小さいことを「水分条件の恒常性」または「恒水性」[homoiohydry; homeohydry]という。反対に、外界の水分条件の変動に強く連動することを「変水性」[poikilohydry]という。
コケ植物や陸生藻類は変水性の傾向が強く、外界が乾燥は植物体の乾燥に直結するが、多くの種が乾燥耐性をもっている(Proctor & al. 2007)。維管束植物はコケ植物より高度な恒水性を備えており、外界の乾燥に対して植物体の乾燥を防ぐことで陸上生活に適応している(シダ植物や被子植物でも、少数だが変水性と乾燥耐性をもつ種が知られている)。しかし、いったん植物体が乾燥するとダメージは大きい。コケ植物は「乾燥に耐える」ことで、維管束植物は「乾燥を防ぐ」ことで陸上の乾燥環境に適応している。
Proctor MCF, Oliver MJ, Wood AJ, Alpert P, Stark LR, Cleavitt NL, Mishler BD. 2007. Desiccation-tolerance in bryophytes: A review. The Bryologist 110:595-621. DOI: 10.1639/0007-2745(2007)110[595:DIBAR]2.0.CO;2
維管束植物が生えることができずコケと地衣だけしか見つからない環境も多い。極端に生育期間が短い高緯度地帯でもコケと地衣が優占し、南極大陸にはセン類約100種・タイ類約25種・地衣類300種以上に対して、種子植物はイネ科コメススキ属のDeschampsia antarctica とナデシコ科のColobanthus quitensis の2種のみが知られていた(現在では、多数の外来種が加わっている)。
維管束植物では、小葉植物・大葉シダ植物・裸子植物・被子植物の全てにおいて樹木という生活型が進化し、森林生態系の出現につながった。古生代デボン紀には、湿潤なところでは森林、乾燥したところでは草原など、様々な生態系が成立し、そこでは、陸上植物と他の生物群との生態的関係が見られた。
陸上植物と他の生物との間に見られる、最も普遍的な関係は、(広い意味での)「食べる―食べられる」の関係だ。陸上における光合成生産の主役として、おびただしい量の植食者[herbivore]や植物病原体[plant pathogen]または植物寄生者[plant parasite]の生活を支える一方、廃棄器官(落葉など)や枯死した植物体を消費して生活する分解者[decomposer]も大量に存在する。
生きた植物を文字通り食べる生物を植食者、内部または表面から栄養を吸収する生物を植物病原体/植物寄生者というが、境界ははっきりしない。また、両者と分解者との境界もあいまいになることがある。
植食[herbivory]の痕跡を残す化石はシルル紀末から出現してデボン紀に増加し、石炭紀の中盤には多様な節足動物、特に昆虫による様々な植食のパターンが見られるようになった。
Labandeira CC 2007. The origin of herbivory on land: Initial patterns of plant tissue consumption by arthropods. Insect Science 14: 259-275 DOI: 10.1111/j.1744-7917.2007.00141.x-i1
菌類・ササラダニ・トビムシなど土壌中の分解者の多くはデボン紀には化石が知られており、植物と分解者の関係はすでに現在とかなり似たものになっていたと考えられる。
分解者との関係が植物にとって(少なくとも直接的には)利益も損害もない(中立)のに対して、植食者・植物病原体との関係は植物に大きな損害(食害・病害)を与える。そのため、植物の方では、トゲや忌避物質、植物毒など色々な防御・回避のしくみが進化し、それに対して植食者・植物病原体の方では解毒能力などの対抗手段が進化した。植食者が植物の進化を促し、植物の進化が植食者の進化を促すというように、複数の生物群が利害関係を通じてからみあいながら多様化することを共進化 [coevolution]という。
デボン紀の陸上植物は、敵対的な「食べる―食べられる」の関係、中立的な分解者との関係、そして、相利的な菌根・共生窒素固定を通じて、おびただしい量の他の生物の生存を支え、現在にいたるまで陸上生態系の基盤となりつづけている。
共生窒素固定: 根粒菌・Frankia属(放線菌)・アナベナ(シアノバクテリア)・ネンジュモ(シアノバクテリア)など、バクテリアの中には大気中の分子窒素(N2 )を植物が利用できるアンモニアに変換する能力を持つものがある。これらのバクテリアが陸上植物の体内に住み、アンモニアを植物に供給することを共生窒素固定といい、コケ・シダ・裸子・被子のそれぞれで一部の種に見られ、土壌の窒素分が不足する環境で優位をもたらすと考えられている。
0-3-3. コケ植物
コケ植物では、胞子体は胞子を散布するためだけに作られる短命な構造で、生活の本拠となる植物体は配偶体だ。配偶体は形状から次の2つのタイプに分けられる。どちらも少数の細胞層でできており、根は持たず、仮根[rhizoid]という細長い細胞の集まりで植物体を基質に繋ぎ止める。
茎葉体: 分岐する軸状の茎に平面的な葉がつく
葉状体[thallus 複数形 thalli]: 全体が平面的で、端が二叉に分かれながら伸長する
コケ植物に含まれる3つのグループは、それぞれ、タイ類・蘚類・ツノゴケ類と呼ばれる。
タイ類 (苔類) [liverwort]: 葉状体を持つものと、扁平で表裏がある(背腹性がある)茎葉体を持つものとがある。
セン類 (蘚類) [moss]: 植物体は茎葉体。
ツノゴケ類 [hornwort]: 葉状体
配偶体は葉状体・茎葉体ともに地表や他物の表面を平面的に広がるのに対して、胞子体は配偶体から上へと突き出すことが多く、胞子が広い範囲に飛散することを可能にしている(タイ類のゼニゴケ類では、配偶体の一部が上に突き出す傘状構造(雌器托・雄器托)をつくり、短い胞子体が傘の下から垂れ下がる)。
葉状体を持つタイ類
左―マキノゴケの葉状体(平面的な暗緑色の広がり)と胞子体(マッチ棒状の直立物)・右―胞子放出後の蒴 トサノゼニゴケ(ゼニゴケ科)。上―雄器托(上の方)と雌器托(下の方)。下左ー葉状体上面(表)・下右―鱗片と仮根がある葉状体下面(裏)。
茎葉体を持つタイ類
オオウロコゴケの植物体・茎葉体・仮根
セン類
コスギゴケの群生・茎葉体・胞子体 上―タチヒダゴケの仮根
ツノゴケ
ナガサキツノゴケ。上―胞子の成熟・散布に至っていない未熟な胞子体をつけた個体。下左―黒緑色の配偶体(葉状体)。下右―葉状体に共生するシアノバクテリア。
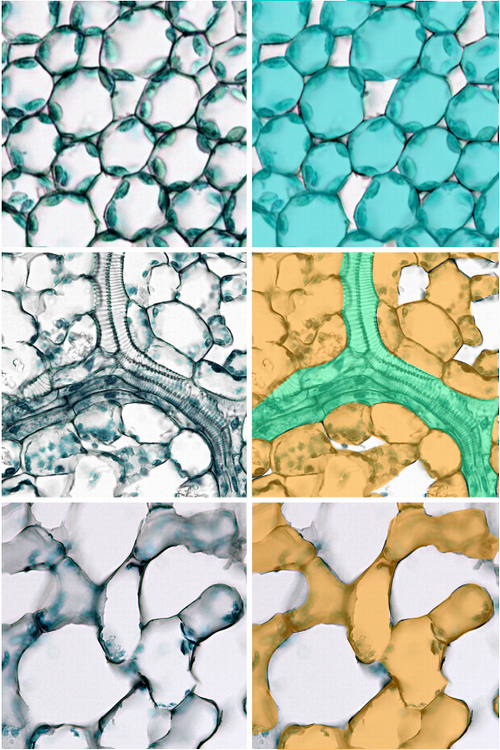
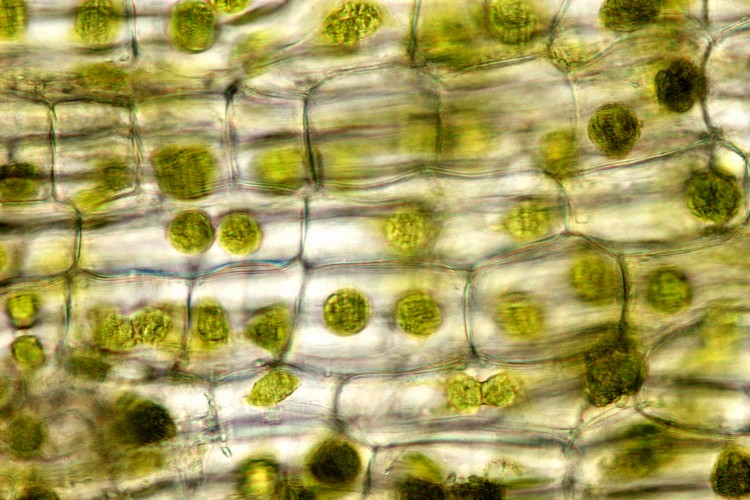
細胞
細胞層が少ないため、コケ植物の葉や葉状体は顕微鏡による細胞の観察が容易だ。典型的なタイ類の細胞は、葉緑体以外に「油体[oil body]」という小粒を含む(数・形状・色は種類によってさまざま)。典型的なツノゴケの細胞は、巨大な葉緑体を1個または2個含む。
コケ植物の細胞。上左―オオウロコゴケ(タイ類)の葉・上右─フルノコゴケ(タイ類)の葉。いずれも細胞内に油体がある。
*)ピレノイド: 葉緑体中にある1個または複数個の顆粒で、多くの藻類とツノゴケに見られる。主成分はルビスコで、CO2固定の場とO2発生の場を分離するしくみと推定されている。緑色植物ではピレノイドが貯蔵デンプンに囲まれていることが多い。
胞子体と胞子散布
胞子体のうち、配偶体とつながっている基底部を「足」、胞子が入っている部分を「蒴」[capsule]という。
タイ類とセン類(一部の群を除く)では足と蒴をつなぐ柄(蒴柄[seta])があり、蒴が裂開して胞子を放出する。タイ類では胞子とともに、弾糸が形成される。弾糸は螺旋状または環状に肥厚した細胞壁による乾湿運動を行い、胞子散布を助ける。セン類の多くでは、蒴は上部が蓋のように外れて、開口部から胞子がこぼれ落ちる(セン類では弾糸は形成されない)。開口部をふちどる歯牙状の構造を蒴歯[peristome]といい、蒴歯の乾湿運動で開口部は開閉する。
ツノゴケ類では蒴柄がなく足に突起状の蒴が直接つき、先端から基部に向かって胞子と弾糸[elator]を形成しつつ、2つに割れ進んで胞子を放出する。ツノゴケの弾糸はタイ類の弾糸のような細胞壁肥厚がなく、「偽弾糸」とよばれることもある。
ツノゴケとセン類の蒴には軸柱[columella]と呼ばれる中心軸があり、蒴の表皮には気孔が分布することがある。タイ類には軸柱・気孔ともみられない。
ナガサキツノゴケの胞子体(左)・胞子体表皮の気孔(右上)・胞子と弾糸(偽弾糸)(右下) 上―ゼニゴケ(タイ類)の胞子体は軸の短いマッチ棒のような形状で、雌器托の傘の下から突き出す。 ハリガネゴケ(セン類)の胞子体。
0-3-4. 小葉植物
小葉植物(ヒカゲノカズラ植物)[Lycophyte]は、ヒカゲノカズラ類・イワヒバ類・ミズニラ類の3つの現生植物群からなる。茎は二叉分岐して原生中心柱をもち、葉は単純な構造で「ミクロフィル(小葉)」[microphyll](phyll=葉)と呼ばれる。
小葉植物(左─ヒカゲノカズラ類のマンネンスギ・右―イワヒバ類のコンテリクラマゴケ)
小葉植物は真葉植物(小葉植物を除いた維管束植物)と姉妹群の関係にある。
0-3-5. 大葉シダ植物
遺伝子から推定された大葉シダ植物の系統関係
マツバラン ハナヤスリなど リュウビンタイなど スギナ・トクサなど ゼンマイなど 薄嚢シダ
大葉シダ植物の中で、マツバラン類(根を持たない/明瞭な葉がない)、トクサ類(スギナ・トクサなど。直立した茎を輪状に取り巻いて小さな三角形の葉がつく)は、飛び抜けて変わったかたちをしており、以前は、シダ植物は、小葉植物・マツバラン類・トクサ類・他のシダ類の4つの系統からなると考えられていた。
上左―マツバラン・上右: ツクシとスギナ(スギナが春先に出す生殖用の茎を「ツクシ」という)・下左―コハシゴシダ・中右─ミゾシダ・下右―マメヅタ
現生の大葉シダ植物の多くの種では、茎は地中や地表・岩表・樹幹にあり、葉だけが空中に広がるが、熱帯・亜熱帯地域ではヤシのような姿の木性シダ(ヘゴ類)が見られる。
古生代石炭紀・ペルム紀の湿地では、小葉植物のリンボク[Lepidodendron]・フウインボク[Sigillaria]や大葉シダ植物トクサ類のロボク[Calamites]などの巨木が「石炭の森」[coal forest]と呼ばれる森林を作っており、枯死後は低酸素条件下で石炭化した。
ヘゴ類(左―ヘゴ・右─マルハチ)
0-3-5. 散布体の進化―胞子から種子
単細胞生物は風や水に流されるだけではなく、鞭毛
多細胞の藻類は、同じ方法をとらず(あるいは、とることができず)、多細胞体の一部を水底などに固定することで固着生活 を送るようになった。そのかわり、有性生殖のため単細胞(卵・精子・受精卵・胞子)になるときに水流に乗って、または鞭毛運動で移動する。
陸上植物のうち、コケ植物とシダ植物では、丈夫な殻で覆われた胞子[spore]を空中に飛散させ(胞子散布)、水の力を借りずに長距離の移動をする。
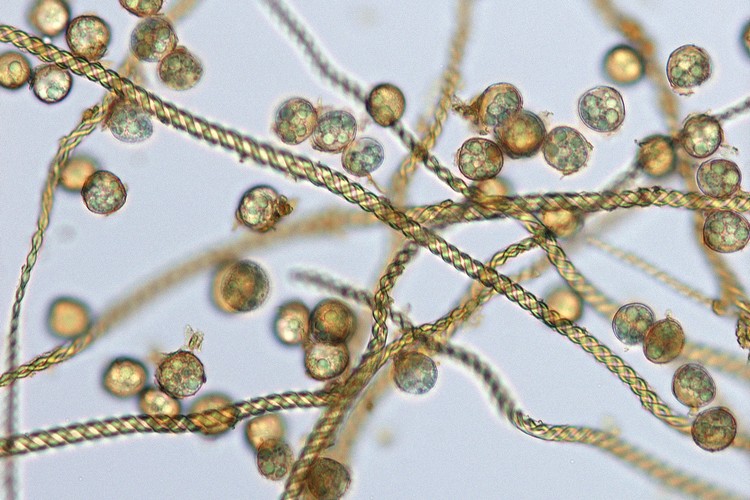
マキノゴケ(コケ植物・タイ類)の胞子の散布。胞子は「弾糸」とよばれる糸状の構造と混じっていて、弾糸が伸びるとともに胞子が溢れだす。 ゼニゴケ(コケ植物・タイ類)の胞子体と裂開 ゼニゴケ(コケ植物・タイ類)の胞子と弾糸 ヤブソテツ(シダ植物)の胞子嚢群の断面
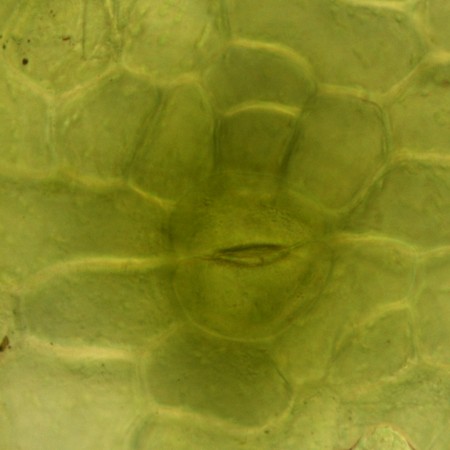
シダ植物では葉の裏や縁に胞子嚢群(ソーラス)がつく。胞子嚢群は、粒状の胞子嚢の集まりで、若いうちは膜(包膜)に覆われていることが多い。熟すると包膜が縮んで胞子嚢が露出する。
ホシダの葉の裏に並んだ丸い胞子嚢群 ホシダの胞子嚢は柄がついたレンズ形で周囲の3/4くらいが縞目のついた環帯に縁取られている。乾燥すると、環帯の収縮運動で胞子を弾き出す。 (videoタグによる動画) (videoタグによる動画) (動画)
胞子は散布先で体細胞分裂をして配偶体(コケ植物では葉状体または茎葉体、シダ植物では前葉体)となる。
マメヅタ(シダ植物ウラボシ科)の培養胞子から発生した細胞塊と前葉体 ミゾシダ(シダ植物メシダ科)の前葉体と前葉体上で発生した胞子体
種子植物の胞子は雄性の小胞子と雌性の大胞子とに分かれ、大胞子に由来する雌性配偶体(胚嚢)は親個体上に留まるため、胞子だけで分布を広げることはできない。
カキノキ(カキノキ科)の種子と縦断面。双葉を持った胚は、茶色の種皮・半透明の胚乳に囲まれている。
種子植物では、受精卵が親個体から養分の供給を受けて細胞分裂し、幼い胞子体=胚となってから親から離れて移動するようになった。
種子 [seed]: 胚と養分を貯える組織がまとめて硬い殻で覆われたもの
種子散布 [seed dispersal]: 種子の移動
種子は胞子より大きくて重いので、移動距離のみを考えると種子散布は胞子散布より不利だ。反面、発生がかなり進んだ状態で移動するので、移動後の発芽・成長ではずっと有利なスタートを切ることができる。また、さまざまな方法で風や動物を使って散布距離を稼ぎ、不利を埋め合わせている。
散布体
単細胞/多細胞
サイズ
散布様式
コケ植物
胞子
単細胞体
小
弾出・飛散
裸子植物
種子
多細胞体
大
多様な散布手段
種子をつける植物(種子植物)はデボン紀に出現し、多数の系統に分岐したが、現在まで残ったのは2つだけだった。1つは被子植物、もう1つは現生の裸子植物の祖先となる系統だ。
0-3-6. 受精様式の進化―精子受精から送受粉+花粉管受精へ
(videoタグによる動画) トサノゼニゴケ(コケ植物・タイ類)の精子と造卵器(動画)
シダ・コケは、鞭毛を持つ精細胞(精子)が雨や地表水の中を移動して受精する(精子受精)。裸子植物・被子植物の共通祖先で、受精は次の2段階で行われるようになった。
送受粉: 精細胞を含む花粉(雄性配偶体)が風や動物などによって胚珠の近くに運ばれる
花粉管受精: 花粉から伸びだした花粉管が精細胞を卵のそばまで運んで受精する
送受粉と花粉管受精の組み合わせによって受精は外部の水に依存しなくなった。また、精細胞の移動距離が格段に長くなって、胞子と比較して短い種子の散布距離を、散布先に同種の生殖個体がいるという限定つきではあるが、埋め合わせた。
受精様式
外部の水
精細胞の移動距離
コケ植物
精子受精
精子が水中を遊泳
必要
短距離
裸子植物
送受粉
様々な媒体が花粉を運搬
不要(水媒を除く)
短距離~長距離
花粉管受精
花粉管が伸長し精細胞を運搬
不要
短距離
ホウセンカの送受粉と花粉管伸長。ハチが花に潜り込んで吸蜜する(左上)と背中に花粉がつく(右上)。他の花に潜り込んで吸蜜するときに背中の花粉が柱頭につく(左下)。柱頭で花粉管は発芽・伸長して(右下)精細胞を卵細胞のところへ運ぶ。 アケビ(アケビ科)の胚珠と花粉管。花粉管の先端は、2枚の種皮に包まれた珠心に入り込む。
脊椎動物の受精でも同じような関係があり、受精が水中で行われる魚類に対し、体内受精を行う爬虫類・鳥類・哺乳類は水に束縛されない生活をしている(両生類には体外受精を行うものと体内受精を行うものとがある)。
裸子植物のイチョウとソテツは精子受精と花粉管受精を併用する例外的な植物で、短い花粉管から放出された精子が卵細胞に達するが、精子が移動するための水分は胚珠から分泌される。
ソテツの胚珠の先端部に入り込んだ花粉は花粉管を伸ばし(左)、精子(右)を放出する
0-3-7. 裸子植物
上左: ソテツ
現生の裸子植物は、3つの系統に分かれる。
ソテツ類
イチョウ
球果類+グネツム類
遺伝子から推定された裸子植物の系統関係
ソテツ類 イチョウ マツ科以外の球果類 マツ科 グネツム類
マツ・スギ・ヒノキなど針葉樹林を構成する樹木を含む球果類が最大のグループだ。球果(種子錐)とは、マツボックリのように種子とそれを守る鱗状の葉が集まってついた器官のことを指す。ただし、イヌマキやイチイのようにマツボックリ状にはならない球果類もある。
スギ(球果類)。枝先に密生する長楕円形の花粉錐(雄花ということもある)から春先に大量の花粉を散らす。花粉は胚珠錐(枝先に集まった小さい灰色がかったふくらみ。雌花ということもある)の中の胚珠につき、花粉管を伸ばして受精する。下の方の大きな緑色のクリのイガのような塊は去年受粉した二年目の種子錐(スギボックリ)。
グネツム類は、グネツム(ツル状で、被子植物の葉によく似た葉をつける)・マオウ(緑色の細い茎が密生)・ウェルウィッチア(2枚の帯状の葉が地を這うように伸び続ける)の3つのグループからなる。植物全体のかたちが他の裸子植物と大きく違うため、独立したグループとして扱うことが多いが、系統的には球果類に含まれる。
左―イチョウの種子(ギンナン)・上―イヌマキの種子。イチョウでは種皮外層が、イヌマキでは種子柄が、目立つ色のみずみずしい可食部となり、動物散布される。
ソテツ類とイチョウの種子は動物散布される。球果類+グネツム類では動物散布と羽根による風散布との両方が、ともに複数回進化し、大形の種子は動物、小形の種子は風によって散布される傾向がある。
Contreras & al. (2017) Evolution of dispersal strategies in conifers: Functional divergenceand convergence in the morphology of diaspores. Perspectives in Plant Ecology, Evolution and Systematics 24:93-117 http://dx.doi.org/10.1016/j.ppees.2016.11.002
Leslie & al. (2017) Variation in seed size is structured by dispersal syndrome and cone morphology in conifers and other nonflowering seed plants. New Phytologist 216:429-437 http://dx.doi.org/10.1111/nph.14456
イチョウ(裸子植物)の胚珠の先端から受粉滴が分泌されている。
花粉を風で運ぶ(風媒)種が多いが、ソテツ類とグネツム類では昆虫による花粉の移動(虫媒)が見られる。多くの裸子植物では、花粉は胚珠の先端から分泌された小さな水滴(受粉滴[pollination drop])に付着し、やがて受粉滴とともに胚珠内に吸い込まれてから花粉管が伸びて先端が卵細胞に達する。
0-3-8. 被子植物の多様化と共進化
アケボノソウ(リンドウ科)の花。茎の先に、萼片×5・花弁×5・雄しべ×5・雌しべ×1(=心皮×2)が規則的に配列している。花弁は複雑な模様に彩られ、2つの蜜腺がついている。
被子植物は、以下のような独特の特徴を持つ。
花 [flower]: 茎の先端に雌しべ・雄しべ・花被片(花びら・萼片など)が一定の配列で密集した有性生殖器官をつくる
裸子植物の有性生殖器官も「花」と呼ぶことがあるが、上記の特徴をもたない
雌しべ: 種子のもとになる胚珠 [ovule]は、雌しべに内蔵されている。花粉は雌しべの先端の特定の部分(柱頭)に付着し、花粉管は雌しべの組織を通って胚珠に到達する。受精した胚珠が種子に変化するのと並行して、雌しべは種子を包む果実 [fruit]へと変化する
重複受精: 精細胞と卵細胞が受精するときに、もう1個の精細胞(1個の花粉粒は、2個の精細胞を持つ)も卵細胞の奥にある中央細胞と受精する
導管を持つ
これらの特徴は、被子植物を他のグループからはっきりと区別し、さらに、被子植物が単系統群
被子植物の現生のさまざまな種の系統関係や地理的分布の変遷は、DNAに基づく系統樹推定と植物化石研究の進展によって解明が進んでいる。現生被子植物の共通祖先は複数のグループに分岐したが、それらのうち、2つのグループが飛び抜けて繁栄し、種数でいうと被子植物の大多数を占めるようになった。
真正双子葉類: キク科・アブラナ科・バラ科・マメ科など
単子葉類: イネ科・ユリ科など
単子葉類の共通祖先では、単子葉性の胚・形成層のない維管束・平行脈を持つ葉などの一連の特徴が進化した(下図では▲で示す)ため、単子葉類は、他の被子植物と形態的にはっきりと区別できるグループとなった。そのため、系統関係が明らかになるまでは、単子葉類以外の被子植物を双子葉類と呼び、被子植物を単子葉類と双子葉類の2つに大別していたが、現在では、真正双子葉類と単子葉類に加えて同じくらい古く分岐した5~6のグループに分ける。そして、真正双子葉類と単子葉類以外を便宜的にまとめて、
基部被子植物
と呼ぶ。
アムボレラ科 基 双
スイレン科など
シキミ科など センリョウ科 モクレン科など マツモ科 単子葉類 ▲ 真正双子葉類 双子葉類 DNAから推定された現生被子植物の系統関係。
コケ・シダ・裸子植物の出現が3億年前以前にさかのぼるのに対して、被子植物の出現は飛び抜けて新しく、最古の化石は中生代中盤で、白亜紀の1億3千万年前から1億年前にかけて急にさまざまな化石が現れ、中生代末~新生代初頭には裸子植物をしのぎ、地球上でもっとも繁栄している生物群の一つとなった。IUCN(国際自然保護連合)のまとめた統計 によると、2020年の見積では、既知の生物種214万種(丸めた数字・以下同じ)のうち被子植物は17%(37万種)を占め、昆虫(105万種・49%)に次ぐ。コケ植物・シダ植物・裸子植物を合わせても3万4千種で被子植物の1割に満たない。
ゲノムの解析からは、裸子植物の祖先と被子植物の祖先の分岐はデボン紀後半~石炭紀前半、被子植物の多様化はジュラ紀中盤に始まって白亜紀前半に急速に進んだと推定されている。その間の2億年あまりの進化の過程は、まだ十分に分かっていない。
既知の生物種数(左)と割合(右): IUCN(2020)による
脊椎動物 無脊椎動物 陸上植物 他
哺乳類 6495 0.3% 昆虫 1053578 49% コケ 21925 1.0% 地衣 17000 0.80% 鳥 11147 0.52% 蛛形類 110615 5.2% シダ 11800 0.55% 真菌 120000 5.6% 爬虫類 11136 0.52% 甲殻類 80604 3.8% 裸子 1113 0.05% 藻類他 23193 1.1% 両棲類 8126 0.38% 軟体 90213 4.2% 被子 369000 17% 魚 35423 1.7% 他 166571 7.8% 小計 72327 3.4% 小計 1501581 70% 小計 403838 19% 小計 160913 7.5% 合計 2137939
小進化の積み重ねで大進化を説明する現代の進化理論の基礎をつくったダーウィン(C. Darwin)にとって、被子植物の急速な多様化の過程は未解明の謎の1つだった。ダーウィンは、植物学者
フッカー(J. D. Hooker)宛ての書簡 で、被子植物が短期間に急速に多様化したことを"an abominable mystery"「忌まわしき謎」と呼んだ。このことにちなんで、アメリカ植物学会誌の2009年1月号は、『種の起源』("On The Origin of Species")の出版(1859)から150周年を記念する
「忌まわしき謎」特集号 として出版された。
このような急速な多様化の要因は完全に解明されたわけではないが、昆虫・鳥類・哺乳類との共進化が大きな原動力の1つとなったと考えられている。
共進化の遺伝子レベルでの基盤として、ゲノムの重複が多様化の直前に起こり、生殖器官の発生を制御する遺伝子の多様化を可能にした、という仮説がある。
被子植物における花から果実への移行の概念図
被子植物では、花で行なわれる送受粉と果実で行なわれる種子散布の両方が、多くの種類において動物との相利関係に依存している。
送粉者―動物媒花
果実食者―糞や貯蔵食として動物に散布される種子
という2つの相利関係は、裸子植物ではごく限られたグループにしか見られないが、被子植物ではふつうに見られる。
ヤマザクラ(バラ科)の花から果実まで。受粉後に花弁、次いで雄しべ・花柱・萼が脱落し、残る子房が成長・成熟する。
送粉動物は花粉食者または花食者、糞や貯蔵食として種子を散布する動物は種子食者または果実食者でもある。送粉や種子散布における相利関係は、「食べる―食べられる」の関係から進化したという説が有力だ。
食べる
花粉食者・花食者
→ 送粉者
相利共生
食べられる
風媒花
動物媒花
食べる
果実食者・種子食者
→ 種子散布者
相利共生
食べられる
非動物散布種子
動物散布種子
ススキ(イネ科)の花と花粉を集める小型のハナバチ。現在でも、風媒花でハチが花粉を集める行動はしばしば観察される。
風媒花でもしばしば花粉を食べたり集めたりする昆虫が観察される。訪花者[flower visitor]≠ポリネーター[pollinator]の例で、植物にとっては損害だ。ただし、風媒花―花粉食者の関係から動物媒花―ポリネーターの関係が起源したのではないか、という説がある。
被子植物の花では、花粉が付着するのは凹凸や分泌液に富んだ雌しべの一部が(柱頭)で、花粉は柱頭の表面で花粉管を伸ばし、花粉管は雌しべに入り込む。胚珠は隔離された子房室内にあるため、花粉食者によるダメージや食害から守られている。このことが、被子植物の進化の初期に、花食者(花粉食者を含む)から送粉者(ポリネーター)へ、風媒花から動物媒花への移行を容易にしたのかも知れない。
ネズミモチ(モクセイ科)の柱頭。花粉粒には3つのスリット(発芽口)があり、スリットから伸び出した花粉管が柱頭の組織に入っていく。
「動物媒花と送粉者」という相利関係が確立した後は、被子植物が動物(特に昆虫)の進化を促し、動物(特に昆虫)が被子植物の進化を促すことで、両者がからみあいながら多様化する共進化の出発点となった。
蜜が露出するヤツデ(ウコギ科)の花は晩秋のハエにとって貴重な食糧源となる ヒガンバナ(ヒガンバナ科)の花で吸蜜しているカラスアゲハ(左)とゲンゲ(レンゲソウ)(マメ科)の花を訪れるセイヨウミツバチ(右)
同じように、種子が果皮に保護されている被子植物では、果実食・種子食という敵対的な関係の一部が「動物に食べられることによる種子散布」という相利関係に移行する敷居が低かった。花の特徴をひもとくのに送粉者の好みや振る舞いの解明が欠かせないように、動物に散布される果実の特徴も散布動物の好みや振る舞いで説明できる部分が大きい。
テキスト目次に戻る ホームに戻る







 このサイクルを「世代交代」といい、減数分裂から受精までを「配偶体世代/n世代/単相世代」、受精から減数分裂までを「胞子体世代/2n世代/複相世代」という。コケ植物では生活の主体は配偶体で、胞子体は配偶体表面にできる分岐しない突起物で、胞子の形成と散布を行う。シダ植物でも胞子体は配偶体(シダの配偶体は「前葉体」と呼ばれる)表面にできるが、配偶体よりも大形になる。種子植物では生活の主体は胞子体で、雌性配偶体(胚嚢)と雄性配偶体(花粉粒)は胞子体の有性生殖器官内部にできる。
このサイクルを「世代交代」といい、減数分裂から受精までを「配偶体世代/n世代/単相世代」、受精から減数分裂までを「胞子体世代/2n世代/複相世代」という。コケ植物では生活の主体は配偶体で、胞子体は配偶体表面にできる分岐しない突起物で、胞子の形成と散布を行う。シダ植物でも胞子体は配偶体(シダの配偶体は「前葉体」と呼ばれる)表面にできるが、配偶体よりも大形になる。種子植物では生活の主体は胞子体で、雌性配偶体(胚嚢)と雄性配偶体(花粉粒)は胞子体の有性生殖器官内部にできる。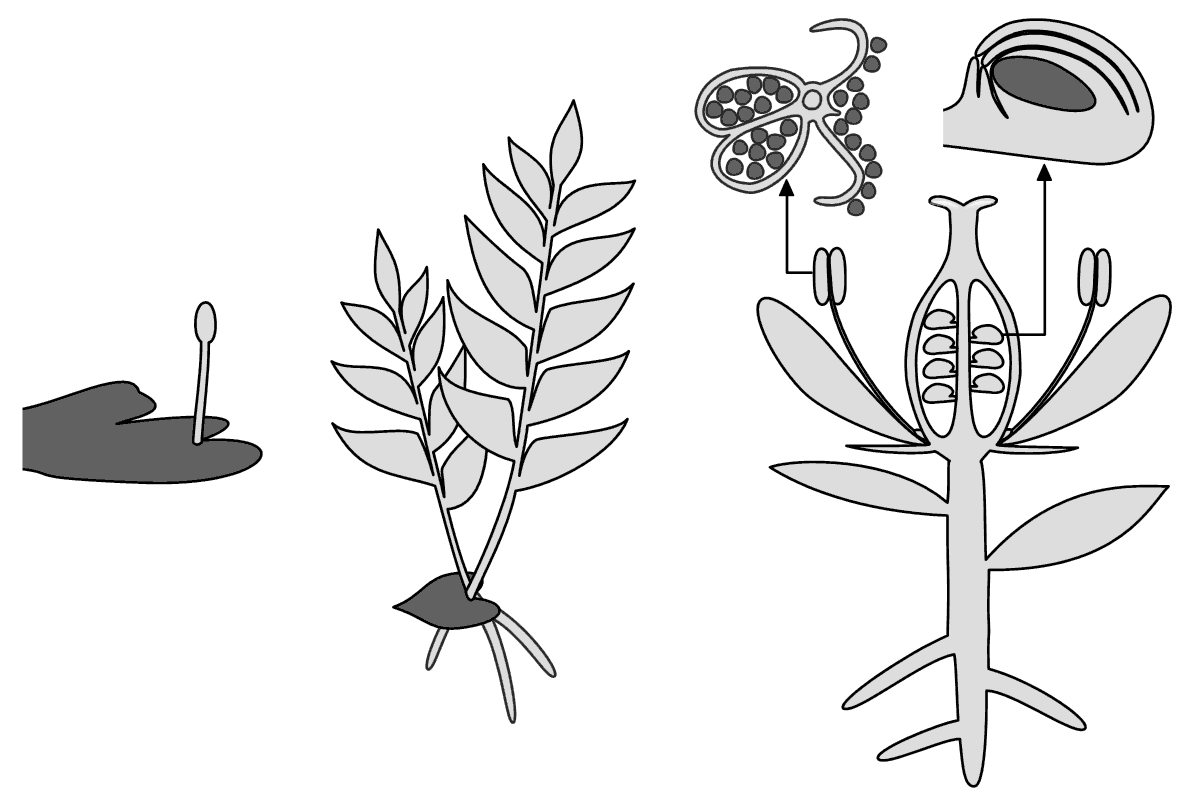















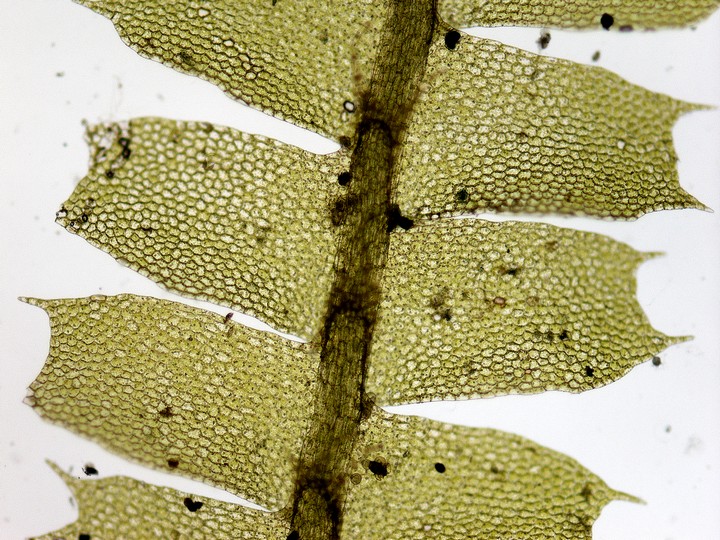










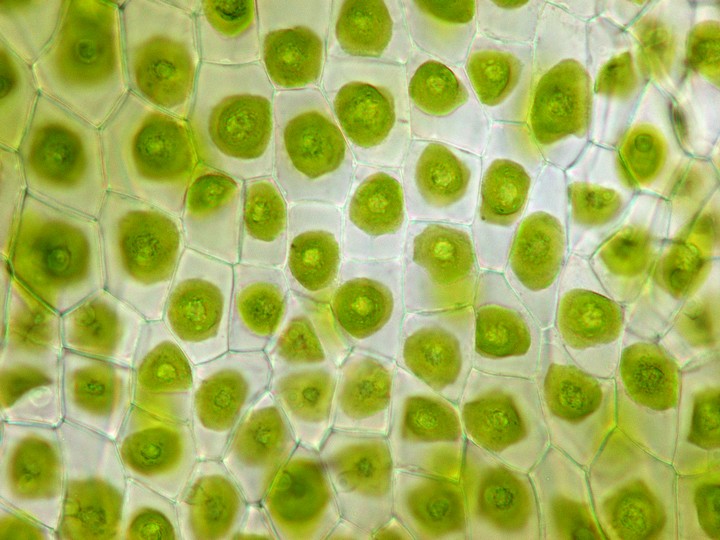

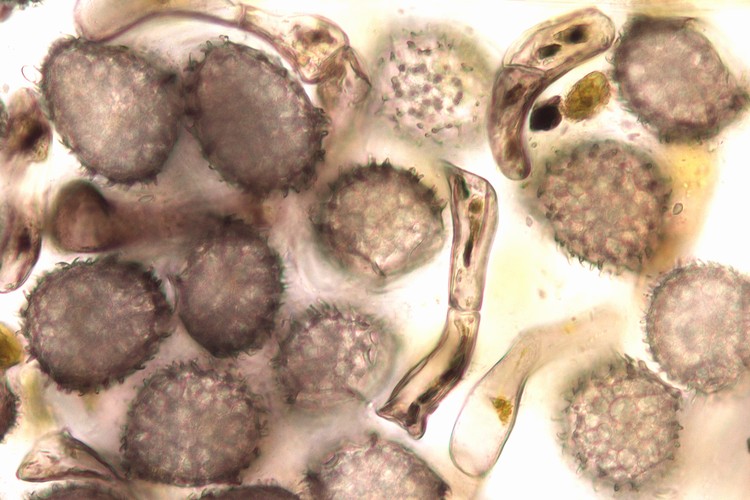

















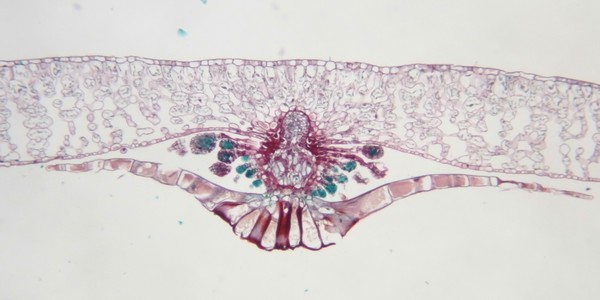 ヤブソテツ(シダ植物)の胞子嚢群の断面
ヤブソテツ(シダ植物)の胞子嚢群の断面
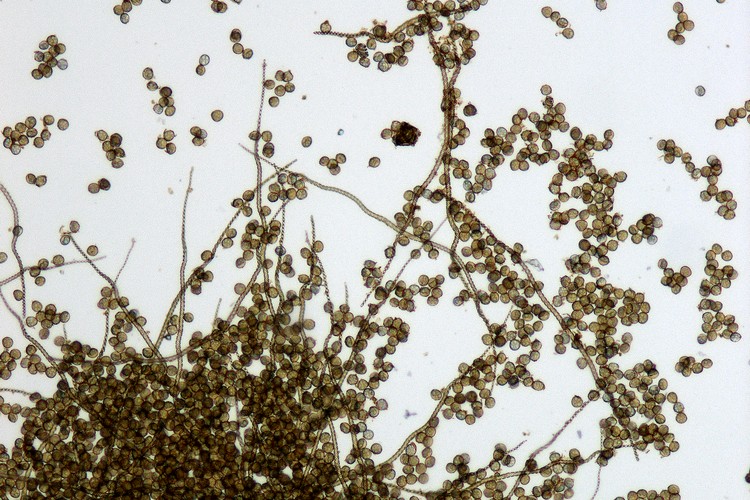
 ホシダの葉の裏に並んだ丸い胞子嚢群
ホシダの葉の裏に並んだ丸い胞子嚢群 ホシダの胞子嚢は柄がついたレンズ形で周囲の3/4くらいが縞目のついた環帯に縁取られている。乾燥すると、環帯の収縮運動で胞子を弾き出す。
ホシダの胞子嚢は柄がついたレンズ形で周囲の3/4くらいが縞目のついた環帯に縁取られている。乾燥すると、環帯の収縮運動で胞子を弾き出す。


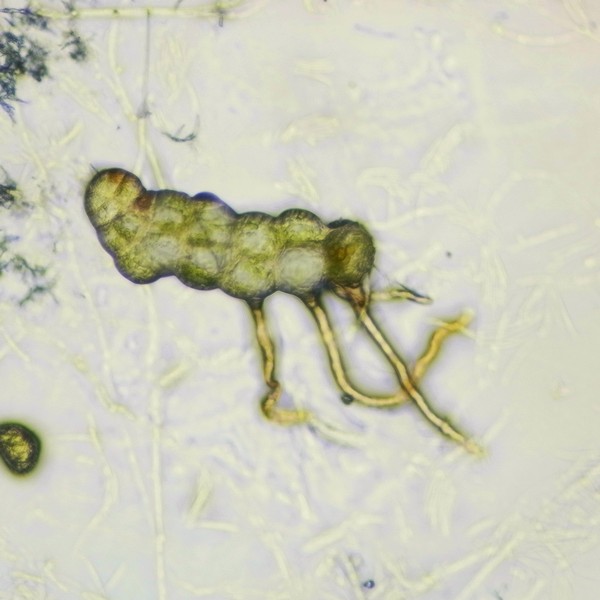
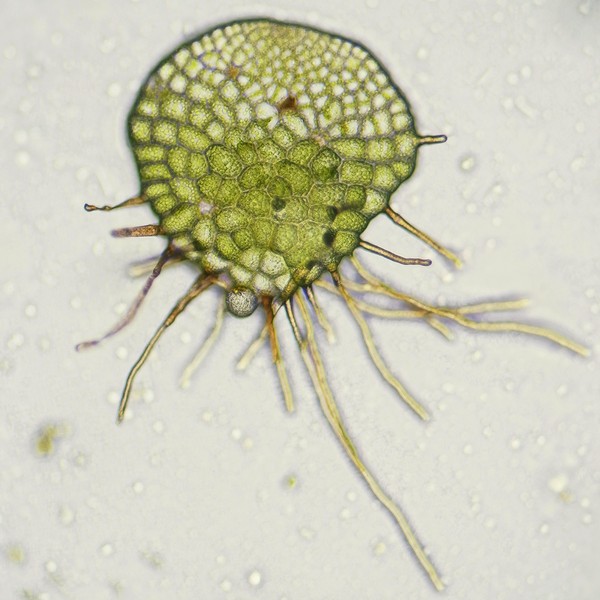 マメヅタ(シダ植物ウラボシ科)の培養胞子から発生した細胞塊と前葉体
マメヅタ(シダ植物ウラボシ科)の培養胞子から発生した細胞塊と前葉体



 ホウセンカの送受粉と花粉管伸長。ハチが花に潜り込んで吸蜜する(左上)と背中に花粉がつく(右上)。他の花に潜り込んで吸蜜するときに背中の花粉が柱頭につく(左下)。柱頭で花粉管は発芽・伸長して(右下)精細胞を卵細胞のところへ運ぶ。
ホウセンカの送受粉と花粉管伸長。ハチが花に潜り込んで吸蜜する(左上)と背中に花粉がつく(右上)。他の花に潜り込んで吸蜜するときに背中の花粉が柱頭につく(左下)。柱頭で花粉管は発芽・伸長して(右下)精細胞を卵細胞のところへ運ぶ。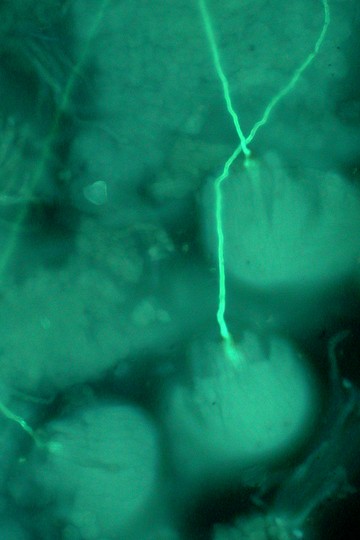

 ソテツの胚珠の先端部に入り込んだ花粉は花粉管を伸ばし(左)、精子(右)を放出する
ソテツの胚珠の先端部に入り込んだ花粉は花粉管を伸ばし(左)、精子(右)を放出する

 上左: ソテツ
上左: ソテツ




 イチョウ(裸子植物)の胚珠の先端から受粉滴が分泌されている。
イチョウ(裸子植物)の胚珠の先端から受粉滴が分泌されている。













