0-2. 陸上植物の出現
地球上の生物は、およそ40億年前に生きていたと推定されるたった一つの祖先から、時間の流れの上に遺伝情報の樹(系統樹
生物進化の歴史の中でも、生命の本格的な上陸=陸上植物の出現は飛び抜けて大きなできごとの1つだ。生命の起源からの長い期間、生物進化の主な舞台は海洋だった。水中(おそらく淡水中)で光合成をしていた生物群の一部が陸上に進出したのは4~5億年前とされる。その子孫は、陸上を主な舞台として進化を続けて多数のグループに分かれ、現在では陸上植物
一部の陸上植物は再び水中に生活の場を移した(水生植物; 第5章)。
地質年代表(単位MYA)
国際地質科学連合国際層序委員会(IUGS-ICS)国際年代層序表 /日本地質学会による邦訳 から作成。陸上進出・維管束の出現・種子の出現と各植物群の盛衰の大まかな順序を横棒で示す(かなりの部分は主観的な評価に基づく)。
冥王代 始生代 原生代 顕生代
4600 4000 2500 541
顕生代
古生代 中生代 新生代
カンブリア紀(Cm)
オルドビス紀(O)
シルル紀(S)
デボン紀(D)
石炭紀(C)
二畳紀(P)
三畳紀(Tr)
ジュラ紀(J)
白亜紀(K)
古第三紀(Pg)
新三+四
541 485 444 419 359 299 252 201 145 66 23
0-2-1. 光合成真核生物の由来
生物系統樹は3つの大グループ(3ドメイン [3 domains])に枝分かれし、各グループがさらに無数のグループに枝分かれする。
バクテリア(細菌) Bacteria
アーキア (古細菌) Archaea
真核生物 Eukarya
真核生物の核遺伝子はアーキアとの共通性が高いがバクテリアと共通する部分もある。ミトコンドリア(実質的に全ての真核生物細胞が持つ)の遺伝子はリケッチア目と、葉緑体(一部の真核生物細胞が持つ)の遺伝子はシアノバクテリアとの共通性が高い。これらのことから、
バクテリアとアーキアが分岐
アーキアが多数の系統に分岐した後、それらの1つが真核生物の祖先となり(エオサイト説[Eocyte hypothesis])
核の進化・αプロテオバクテリアの細胞内共生によるミトコンドリアの獲得
単細胞真核生物の1系統がシアノバクテリアの細胞内共生によって葉緑体を獲得(単細胞光合成真核生物の起源)
真核生物の複数の系統で多細胞体制が進化
という順序が推定されている。
αプロテオバクテリア[Alphaproteobacteria]: 多数の種を含むバクテリアのグループで、細胞内寄生体/共生体としては発疹チフスなどの病原体リケッチア[Rickettsia]や節足動物の細胞内に寄生するボルバキア[Wolbachia pipientis]、根粒バクテリア[Rhizobium]など、非寄生の(自由生活性の)種としてはあらゆる地域の海水中に大量に存在するSAR11[Pelagibacter ubique]や一部の光合成バクテリアなどを含む。
シアノバクテリア[cyanobacteria]: アオコ・イシクラゲなどを含む光合成バクテリアのグループ。「藍藻」[blue algae]と呼ばれることもある。アオコとイシクラゲの細胞群体
葉緑体をもつ生物=光合成真核生物は、生物系統樹の様々な枝に飛び石状に分布している。このことは、以下の様に説明されている。
シアノバクテリアの細胞内共生(一次共生)による葉緑体の獲得は(後述のビンカムリ類を除いて)単一の出来事であった。一次共生によって葉緑体を獲得した共通祖先に由来する生物群を一次植物といい、生物系統樹上で1つのまとまった枝(アーケプラスチダ)をつくる。
一次植物の細胞内共生(二次共生)による葉緑体の獲得が様々なグループで複数回起こった。二次共生による葉緑体をもつ生物群を二次植物といい、生物系統樹上の様々な枝に散在する。
いったん獲得した葉緑体の消失も様々なグループで複数回起こった。
葉緑体獲得の模式図。
Nozaki H, Maruyama S, Matsuzaki M, Nakada T, Kato S, Misawa K. 2009. Phylogenetic positions of Glaucophyta, green plants (Archaeplastida) and Haptophyta (Chromalveolata) as deduced from slowly evolving nuclear genes. Mol. Phylogenet. Evol. 53 (3): 872-80. doi:10.1016/j.ympev.2009.08.015
Burki F, Shalchian-Tabrizi K, Pawlowski J. 2008. Phylogenomics reveals a new 'megagroup' including most photosynthetic eukaryotes. Biology Letters 4(4): 366-369. doi:10.1098/rsbl.2008.0224
藻類
光合成をする真核生物=葉緑体を持つ真核生物のうち、陸上植物以外を総称して藻類[algae]という。陸上植物の進化と生活の場が陸上であるのに対し、藻類は水中を主な舞台として進化し、現在も水中を主な生活の場としているり、水中を主な舞台として進化し、現在も水中を主な生活の場としている。水中で生活する陸上植物がある(水生植物)ように、陸上の様々な環境(土壌や地表、岩表、樹皮など)で生活する藻類もあり、気生藻(類)ないし陸生藻(類)と呼ばれる。
シアノバクテリアを(便宜上)藻類に含める場合は「藍藻」と呼ぶ。
藻類は単細胞・細胞群体・多細胞など様々な体制のものがあり、系統的な位置もきわめて多岐にわたる。
一次植物に含まれるグループには、陸上植物・ストレプト藻・緑藻を含む「緑色植物」と呼ばれるグループ、紅藻(アサクサノリ・テングサ・フノリ類など)、灰色(かいしょく)藻などがある。
一次植物の例。左―オオヒゲマワリ(緑藻)・右―フクロフノリ(紅藻)
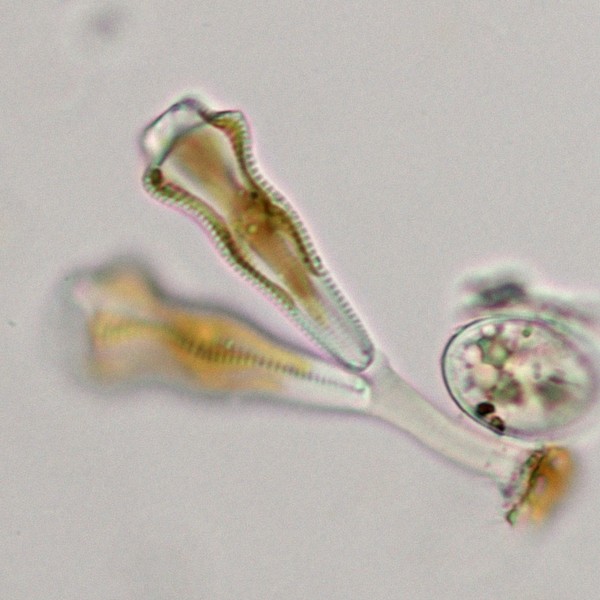
二次植物は、生物のさまざまな系統群に分散している。例えば、卵菌(ミズカビ・べと病菌・白サビ病菌・ジャガイモ疫病菌など)と比較的近い褐藻(コンブ・ワカメなど)・珪藻は、紅藻の二次共生による葉緑体をもつ。また、トリパノソーマ(病原体の1つ)と同じグループに属するミドリムシ(ユーグレナ)類は、緑色植物の二次共生による葉緑体をもつ。繊毛虫(ゾウリムシなど)・マラリア原虫と比較的近い渦鞭毛藻では、祖先系統が紅藻の二次共生によって葉緑体を獲得したが、葉緑体の退化や二次共生で獲得した葉緑体をもつ単細胞生物の細胞内共生(三次共生)、さらに別々の藻類による複数回の二次共生などがあり、非常に複雑な葉緑体の進化があったことが知られている。
二次植物の例。左―ホンダワラ類の1種(褐藻)・上―スジタルケイソウの1種(珪藻) 二次植物の例。左―ツノオビムシの1種(渦鞭毛藻)・右―ミドリムシ(ユーグレナ)の1種
もう一つの一次共生
現生の光合成真核生物の葉緑体の元になった一次共生はただ1回の出来事だったと推定されている。例外は、ケルコゾアに属する単細胞真核生物ビンカムリ Paulinella chromatophora (とその近縁種)だ。ビンカムリの葉緑体は、
他の葉緑体とは違う系統のシアノバクテリアと遺伝子が似ている
他の葉緑体と比べて遺伝子の核への移行の程度が低い(葉緑体やミトコンドリアでは、必要な遺伝子のかなりの部分が核のDNAに移行しており、遺伝子レベルで独立性を失っている)
等の特徴をもち、他の生物の葉緑体とは別に、ずっと新しく起源したと推定されている(例えば、Marin B, Nowack EC, Melkonian M. 2005. A plastid in the making: evidence for a second primary endosymbiosis. Protist 156:425-432. https://doi.org/10.1016/j.protis.2005.09.001 )。
独自の起源をもつ葉緑体は、今後他に見つかる可能性もある。絶滅によって痕跡が失われた一次共生が生物進化の過程であったとしても不思議ではない。
「植物」の範囲
「植物」が指す範囲は歴史的に変化してきた。
かつては、運動能力がないあらゆる生物を指した。1900年代初頭の植物学の教科書を見ると、植物界は「顕花植物」(種子植物)と「隠花植物」に二分され、隠花植物は、シダ・コケに加えて菌類・バクテリアも含まれている。
光合成能力(=葉緑体をもつこと)を中心に植物が定義されるようになり、菌類・バクテリアは除外されて、植物=陸上植物+藻類となったが、二次共生による葉緑体の起源が明らかになり、葉緑体をもつことと単一起源を持つグループとが相容れないことがはっきりした。
現在では、系統関係を重視して定義することが増えている。
植物=一次植物(=アーケプラスチダ)
植物=緑色植物
植物=陸上植物のみ(高等学校生物で採用している定義)
と、陸上植物を含むことは共通で、範囲づけに広い狭いがある。
植物=光合成真核生物(陸上植物と藻類の総称)とする伝統的な定義やその流れを汲む用語は、日常生活でしばしば使われている(例: 植物プランクトン)だけでなく、専門用語にも少なからず残存している(例: 二次植物)。
0-2-2. 陸上植物の出現
カンブリア紀の動物群の爆発的な多様化によって、水界(海洋・河川・湖沼)では現在のかたちに近い生態系が成り立っていた。しかし、海洋の大部分を占める外洋では、光の当たる表層では養分が不足、養分が溜まる深層では光が不足する。大陸に近い浅海や河川、浅い湖沼にはその両方があるが、面積は限られている。
それに引き換え陸上では、養分を含む土壌に十分な光が降り注ぐ。光合成をする生物にとっては、外洋にない好条件で、しかも、浅海・河川・湖沼とは比べものにならないほど広大だ。
陸上には、生物にとって水界にない悪条件もあり、バクテリアなどの微生物が地表や土壌中で生活していたに過ぎなかった。
強い紫外線
乾燥
寒暖の差
初期の陸上植物はいくつかの特徴によってこれらの困難をしのいだ。
外殻に覆われ空中を飛散できる胞子
紫外線を吸収する色素群(フラボノイドなど)
細胞が立体的に結合した多細胞体
植物体表面を覆う透明層(クチクラ)
陸上植物と推定される最初期の化石群は胞子で、オルドビス紀の地層から発見されている。シルル紀以降の地層では、維管束や気孔の化石も加わる。
陸上植物は多数のグループに分岐し、その中から、さらに新たな特徴を獲得し、より複雑かつ立体的で大型の身体を作るようになったものが出現した。
根・維管束・クチクラ・気孔からなる吸水とガス交換のシステム(維管束植物)
送受粉と花粉管受精(種子植物)
種子散布(種子植物)
これらの過程を通じて、陸上植物は他の陸生生物とさまざまな相利関係を結び、陸上生態系はより立体的で多様性の高いものに変化していった。
菌根: 根―菌類
動物送粉: 花―動物(特に昆虫と鳥類)
動物被食型種子散布: 種子+α―動物(特に鳥類とサル)
緑色植物
陸上植物と近縁な藻類を含む「緑色植物」[Viridiplantae]は、以下のように分類される(主なグループのみ)。
ストレプト植物
陸上植物
接合藻(アオミドロ・ツヅミモ・ミカヅキモなど)
広義のシャジクモ植物
広義の緑藻
コレオケーテ(サヤゲモ類)
車軸藻(シャジクモ類)
緑藻植物
緑藻(ボルボックス・クラミドモナス・クンショウモ・イカダモなど)
トレボキシア藻(クロレラなど)
アオサ藻(アオサ・ヒトエグサなど)
単に「緑藻」というとき、狭義の緑藻を指す場合、緑藻植物を指す場合、陸上植物以外の緑色植物を指す場合がある。
陸上植物の祖先は、広義のシャジクモ植物(いずれも淡水生)に近い生物群に属していたと推定される。
上: シャジクモ。軸途中の関節状のところに有性生殖器官をもつ。苞に囲まれてねじれた外殻を持つのが雌性生殖器官(生卵器)で、卵細胞が1個入っている。生卵器の下方につく球状の雄性生殖器官(造精器)では多数の精子が作られ、放出される。受精は生卵器内で起こる。
シャジクモ・コレオケーテは、陸上植物を連想させる複雑な体制をもち、成長様式や有性生殖、細胞分裂においても陸上植物との共通点が多く、以前は陸上植物と最近縁と考えられていた。
成長様式: 軸先端での細胞分裂による成長
有性生殖: 造精器から放出された精子が生卵器の卵細胞に受精
細胞分裂: 細胞板形成による細胞質分裂; 原形質連絡の形成
しかし、DNA塩基配列に基づく系統推定の進展によって、接合藻が陸上植物に最近縁である(陸上植物と最後に分岐した)ことが有力視されるようになった。
接合藻の例
接合藻の多くの種はアオミドロやツヅミモ、ミカヅキモにように淡水生だが、準陸生の種も含まれており、湿った地表や岩上に生育し、ある程度の乾燥耐性[Desiccation tolerance; DT]をもつ準陸生の種も含まれている。接合藻が陸上植物の最近縁であれば、準陸生の接合藻がもつ乾燥耐性が陸上植物の祖先の陸生生活へ適応に貢献したという想定が可能だ。この想定を裏づけるように、乾燥などのストレス耐性に関与する遺伝子(GRAS・PYR/PYL/RCAR)が、準陸生接合藻と陸上植物のゲノムから見出され(シャジクモ等にはない)、近似の遺伝子をもつ一部の土壌生バクテリアから遺伝子水平移動によってもたらされたと推定された。
Cheng et al. (2019) Genomes of subaerial Zygnematophyceae provide insights into land plant evolution. Cell 179, 1057–1067 https://doi.org/10.1016/j.cell.2019.10.019
フラボノイドとリグニンの生合成
フラボノイド : フラバンを基本骨格とするきわめて多様な化合物群で、ほとんどの陸上植物に存在する一方、藻類では発見されていない。紫外線吸収や花色色素などさまざまな役割を担う。
左: アカメガシワ(トウダイグサ科)の展開中の新葉。フラボノイドに属する色素群アントシアニンによって新葉が赤く着色する植物は広く見られ、紫外線から防護する役割をもつとされる。
リグニン : モノリグノール(p-クマリルアルコールなど)が重合した複雑な立体網目構造をもつ高分子。維管束をもつ陸上植物に見られ、道管などに見られる堅い細胞壁の主要な成分となる(藻類の紅藻の一部でもリグニンが見出され、維管束植物とは別の起源をもつと考えられている)。
カボチャ茎横断面の厚壁組織とクスノキ導管要素の細胞壁
フェニルアラニンを脱アミノ化する酵素フェニルアラニンアンモニアリアーゼ(PAL)は、陸上植物・菌類・バクテリアに存在するがシャジクモ等の藻類では未発見で、陸上植物の祖先がバクテリアから遺伝子水平移動[Horizontal gene transfer (HGT) または Lateral gene transfer; LGT]によって獲得したという推定がある。
PALとGRAS・PYR/PYL/RCARの例に見られるように、全ゲノム解析に基づく研究の進展によって、先行して上陸していたバクテリアからの遺伝子水平移動が陸上植物の出現に果たした役割が重視されつつある。
テキスト目次に戻る ホームに戻る



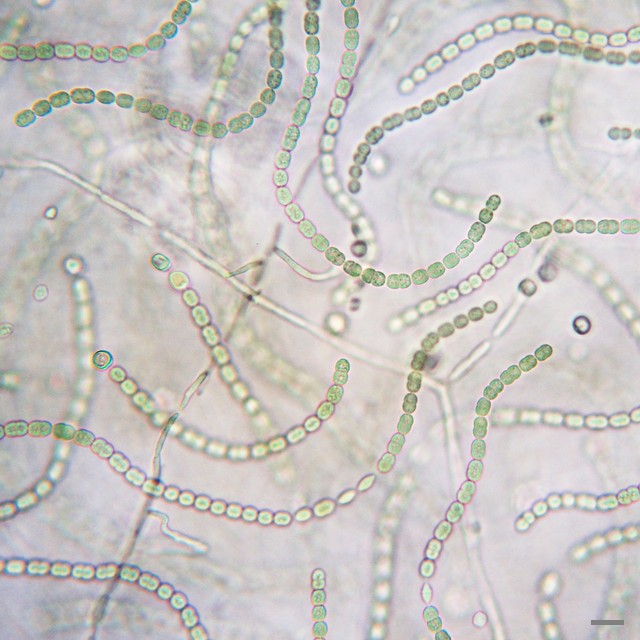
 左―ミドリゾウリムシ(繊毛虫類)にはクロレラが内部共生している
左―ミドリゾウリムシ(繊毛虫類)にはクロレラが内部共生している

 葉緑体獲得の模式図。
葉緑体獲得の模式図。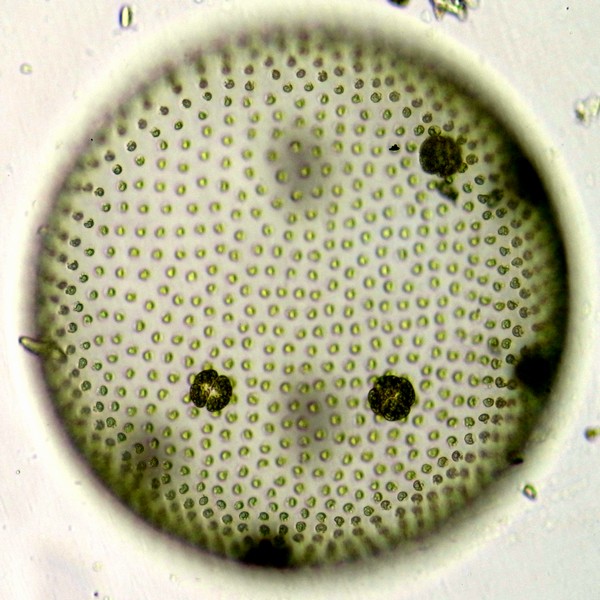






 上: シャジクモ。軸途中の関節状のところに有性生殖器官をもつ。苞に囲まれてねじれた外殻を持つのが雌性生殖器官(生卵器)で、卵細胞が1個入っている。生卵器の下方につく球状の雄性生殖器官(造精器)では多数の精子が作られ、放出される。受精は生卵器内で起こる。
上: シャジクモ。軸途中の関節状のところに有性生殖器官をもつ。苞に囲まれてねじれた外殻を持つのが雌性生殖器官(生卵器)で、卵細胞が1個入っている。生卵器の下方につく球状の雄性生殖器官(造精器)では多数の精子が作られ、放出される。受精は生卵器内で起こる。








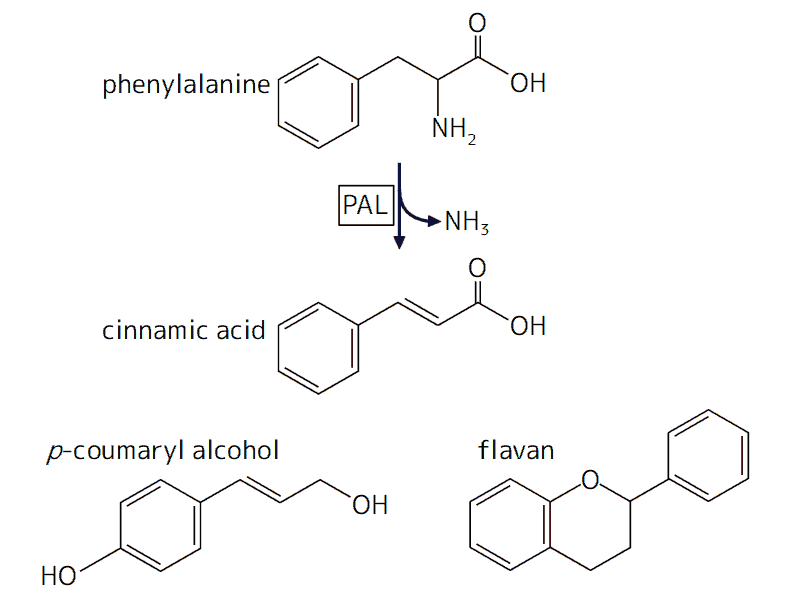 ベンゼン環をもつアミノ酸フェニルアラニンが脱アミノ化して生じた桂皮酸(シナモン酸)は、フラボノイドやリグニンなど、陸上への進出や適応の鍵となる多様な物質の合成の出発点となる。
ベンゼン環をもつアミノ酸フェニルアラニンが脱アミノ化して生じた桂皮酸(シナモン酸)は、フラボノイドやリグニンなど、陸上への進出や適応の鍵となる多様な物質の合成の出発点となる。