6-1. 花を構成する要素
6~8章で扱う花・花序・果実は有性生殖に直接かかわる「有性生殖器官」で、2~5章で扱った「栄養器官」と対置される。
6-1-1. 「花」=被子植物の有性生殖器官
被子植物の特徴の一つは、有性生殖をするときシュートの先端が「花」[flower]と呼ばれる器官をつくることだ。
日常感覚における「花」と生物学における「花」
日常使っている表現の多くが、「花の色」「花が咲く」「花が散る/しぼむ」など「花」と花弁を同一視している。このような感性を反映してか、「花」は、文化的伝統において「美しいもの」「目立つもの」「栄えているもの」かつ「はかない(短命な)もの」「無常なもの」「移ろうもの」として扱われてきた。
古事記・日本書紀では、ニニギノミコトは「美・繁栄・短命」を象徴するコノハナサクヤヒメに求婚して「醜・耐久・永遠」を象徴するイワナガヒメを拒絶し、死の宿命を負うことになる(世界中にある「死の起源神話」(Origin-of-death myth; ODM)の一類型)。
万葉集収録の大友家持の2首「咲く花は 移ろふ時あり あしひきの 山菅の根し 長くはありけり(20/4484)」「八千種の 花は移ろふ 常盤なる 松のさ枝を 我れは結ばな(20/4501)」は短命な花と長久な山菅の根/松の枝とを対比し、古今和歌集巻二(春・下)では桜の花の儚さを詠む歌が延々と続く。例えば、「花の木も 今は掘り植ゑじ 春た立てば 移ろふ色に 人倣ひけり」(素性法師)。
十七世紀の西欧では、花・宝飾品・楽器・若人と骸骨・死体とを組み合わせて「富や繁栄、快楽の空しさ/儚さ」という人生訓を暗示する「ヴァニタス」[Vanitas]と銘打たれた静物画がさかんに描かれた。
生物学における「花」は殆ど正反対の性格をもつ: 花の不可欠な「真の主役」は雌しべと雄しべに他ならない(花弁がない花は少しも珍しくない)。花は(1)送受粉・(2)果実になる、の2つの役割を通じて生命の連続性を担う器官で、進化を通じて獲得された巧みで逞しいしくみが凝縮している。
花では、中心(茎頂に当たる)から
- 雌しべ[pistil; gynoecium]
- 雄しべ[stamen; androecium]
- 花被片(かひへん)[tepal] (花弁と萼片)
が同心円状からせん状に並んでいる。後で述べるようにこの通りでない花も無数にあるが、部品の一部が退化(欠失)したり、複数の部品が一体化(合着)した結果と考えることができる。
ホンゴウソウ科のごく小さい単子葉の腐生植物、ラカンドニア・スキスマチカ Lacandonia schismatica は雄しべと雌しべの順序が「逆転」する唯一の例外として有名だ。雄しべが花の中央を占め、その周りを雌しべが取り巻いている(近縁種の Triuris brevistylis でも低頻度で逆転が見られることがある)。1985年にメキシコの熱帯雨林で発見されて大きな話題となった。


サクラ(カラミザクラ)の花を上と下から見たところ。1個の雌しべ・約40本の雄しべ・5枚の花弁・5枚の萼片(基部はつながって筒になっている)がある。
下は縦断面。萼片の基部はつながって筒になり、筒の中に子房が納まる。

| 単独での呼び方 |
花1つ分まとめて
呼ぶとき |
| 花被片 |
萼片 |
花被 |
萼 |
| 花弁(花びら) |
花冠 |
| 雄しべ |
雄しべ/雄しべ群 |
| 雌しべ |
雄しべ/雌しべ群 |
花被片は、内側の花弁(花びら)[petal]と外側の萼片(がくへん)[sepal]の2種類に分けられることが多い。また、1つの花の花被片をまとめて「花被」[perianth]と呼ぶ。同じように、1つの花の花弁をまとめて「花冠」[corolla]、1つの花の萼片をまとめて「萼」[calyx]と呼ぶ。


アサガオ(ヒルガオ科)の花冠とホウセンカ(ツリフネソウ科)の花被
このような「まとめる呼び方」は、アサガオのように花弁(5つ)が融合・一体化している場合(「花冠はラッパ状」)や、ホウセンカのように花弁(3つ)と萼片(3つ)が組み合わさって特徴的な形状を作っているとき(「花被は覆いのあるちりとり形」)に便利だ。
雄しべ・雌しべでも同様に「雄しべ群」「雌しべ群」とまとめることがあるが、文脈で分かる場合は「雄しべ」「雌しべ」で済ませることも多い。当然ながら、雌しべが花に1つのときは必ず「雌しべ」を使う。
次の項で述べるように、花はシュートの先端、萼片・花弁・雄しべは、それぞれがシュートについている葉に相当する。雌しべは1枚の葉に相当する場合と、複数の葉の集合体である場合とがある(あとでもう少し詳しく解説する)。






花の模式図。萼片と花被片がはっきりと区別できるタイプの花を例に取っている。
1―花柄(花梗)、2―花托(花床)、3―萼片(がく片)、4―花弁(花びら)、5―蜜腺、6―雄しべ(雄蕊)、7―雌しべ(雌蕊)。
蜜腺は花蜜を分泌する組織で、図のように雄しべ・雌しべのつけねに独立した突起としてできる場合もあるが、他のさまざまな部位にできることもある。
胞子嚢穂
胞子をつける葉[sporophylls]または枝[sporangiophores]がシュートの先端に密集した構造を胞子嚢穂[strobilus 複 strobili]といい、維管束植物の現生種・化石種のさまざまなグループに見られる。グループや雌雄によって構造は大きく異なる。
-
小葉植物のヒカゲノカズラ・大葉シダ植物のスギナなどに見られる「狭義の胞子嚢穂」
-
裸子植物の花粉錐[pollen cone]・胚珠錐(種子錐)[ovule cone (seed cone)]: 「球花」「球果」と呼ぶこともある。
-
被子植物の花
左―ヒカゲノカズラの胞子嚢穂、上中―スギナの胞子嚢穂、上右―カラマツの花粉錐、下右―カラマツの胚珠錐
裸子植物の花粉錐は各グループ間で比較的よく似ており、スギナやヒカゲノカズラの胞子嚢穂とも共通性が高い。これに対して胚珠錐の構成はグループ間の違いが非常に大きい。
-
ソテツ: シュートの先端にゆるく集まった羽状の大胞子葉[megasporophyll]のへりに胚珠がつく
-
イチョウ: 細い枝の先が二叉に分かれ、それぞれに1個ずつの胚珠がつく
-
マツ・スギ・ヒノキなどの種子錐(球果)は、中軸に二枚重ねの鱗片(外側の苞鱗・内側の種鱗)がびっしりとついた構造で、種鱗の内側に1対の胚珠がつく。苞鱗は中軸につく葉、種鱗は苞鱗の腋のシュートが変形した構造と解釈されることが多い。
花粉錐・胚珠錐と被子植物の花との関係は不明な部分が大きいため、裸子植物には「花」を使わず、被子植物に限定する方がすっきりと定義できる。

 クロマツ(マツ科)の4月(送粉期)と6月。春に新しく伸びたシュートの先端に少数の胚珠錐、基部に多数の花粉錐がつく。胚珠錐は次の年の秋に熟して種子錐となり、種子を出し、多くはその翌年まで枝についたまま残る。
クロマツ(マツ科)の4月(送粉期)と6月。春に新しく伸びたシュートの先端に少数の胚珠錐、基部に多数の花粉錐がつく。胚珠錐は次の年の秋に熟して種子錐となり、種子を出し、多くはその翌年まで枝についたまま残る。
日本の中高理科では、strobilus・花粉錐・胚珠錐などの用語を避けて、裸子植物の花粉錐を「雄花」、胚珠錐を「雌花」と呼ぶ(胚珠錐を「雌花の集まり」としていた時期もある)。種鱗を鱗状の枝と見なせば、種鱗=「雌花」の方が被子植物の花により適合するが、現在では分かりやすさを優先して胚珠錐=雌花とする。なお、花粉錐(雄花)につく花粉嚢(花粉袋)を「葯」としていた時期もあるが、現在は「葯」は被子植物の雄しべだけに使う。
花序
花は、単独で枝につく(単生)こともあるが、多くの種では若い枝に集まってつく。このような枝のことを花序[inflorescence]と呼ぶ。
果実が集まってついた枝、つまり、花が終わって果実がつく状態となった花序を果序[infructescence]という(花序と発音が同じで、紛らわしい)。
 セイヨウアブラナの花序。左―横から見たところ、下―上から見たところ(拡大)。花は中心軸上につき、下(基部)の花から先に咲く。軸の先端付近ではつぼみが密集している。開花の進行とともに軸が伸びるため、軸の下の方で咲き終わった花が間隔を置いてつくようになる。
セイヨウアブラナの花序。左―横から見たところ、下―上から見たところ(拡大)。花は中心軸上につき、下(基部)の花から先に咲く。軸の先端付近ではつぼみが密集している。開花の進行とともに軸が伸びるため、軸の下の方で咲き終わった花が間隔を置いてつくようになる。

花序は、ふつうの枝と比べて、以下のような点で違っていることが多い。
-
他の枝よりも突き出している
-
葉はふつうの葉よりも小さくて単純な形になっている
-
枝が密に分かれている
これらの特徴は、花序についている花が自身の葉や枝に隠されたり、紛れたりする可能性を低くしている。
花序のようす(分岐型・発生パターン・開花順序・全形など)は、グループによって異なり、送受粉のあり方や、まれには種子散布の様式とも深く関わっている。(→花序の開花順序)
花の(究極的な)はたらき・構成・発生の遺伝的背景は、被子植物を通じて共通している。一方、形・色・サイズは、きわめて多様だ。






被子植物の花の多様性
6-1-2. 有性生殖の基本器官 (1)雄しべ
 雄しべ・葯(横断面)・花粉の模式図。グループによってさまざまで、わりとよく見られるタイプに似せて描いたもの。
雄しべ・葯(横断面)・花粉の模式図。グループによってさまざまで、わりとよく見られるタイプに似せて描いたもの。
A―雄しべ
B―葯の断面(左半は裂開前、右半は裂開後)
C―葯壁の断面
D・E―花粉粒(D―三溝粒、E―単口粒)。
1―葯、2―半葯、3―花糸、4―葯隔、5―葯壁、6―花粉粒、7―発芽口、8―花粉壁。
雄しべは、花粉袋(葯 やく[anther])と、葯をつける柄の部分(花糸[filament])からなっている。葯は多数の花粉粒[pollen grain]とそれを包む皮からできている。花粉粒の集まりを花粉[pollen 不可算名詞]という(花粉粒を「花粉」と呼ぶこともあるが、本当は間違った使い方)。


セイヨウアブラナ(アブラナ科)のつぼみと葯の横断面。花弁が開く直前に葯が花粉を出す。葯室は4つ。葯壁が裂けて花粉の出口2ヶ所ができている。
葯は、軸(花糸の先端にあたり、維管束が通っている)の両側にある2つの半葯[theca 複-thecae]でできている。半葯には1つか2つの葯室(花粉嚢)があるので、葯には合計2つか4つの葯室がある。葯室の壁(葯壁[anther wall])が裂けて花粉が葯から外に出られるようになる。セイヨウカラシナのように、葯室が4つでも、花粉が出る裂け目はふつう2つしかない。


アオカズラ(アワブキ科)の花と葯の横断面。葯室は2つ。
 ナガバタチツボスミレ(スミレ科)の葯壁。左の写真は垂直に切った断面で、右側が内側(花粉がある方)にあたる。最も発達した中央の細胞層では、内側の細胞壁が肥厚しており、外側の細胞壁は薄いままだ。このような構造は、やはり裂けて中身を吐き出すシダの胞子嚢にも見られる。
ナガバタチツボスミレ(スミレ科)の葯壁。左の写真は垂直に切った断面で、右側が内側(花粉がある方)にあたる。最も発達した中央の細胞層では、内側の細胞壁が肥厚しており、外側の細胞壁は薄いままだ。このような構造は、やはり裂けて中身を吐き出すシダの胞子嚢にも見られる。
葯から花粉が外に出られるようになることを花粉表出(仮訳)[pollen presentation]という。葯壁に縦に裂け目が入って花粉が露出するケース(縦裂開)が最も多数を占める。

ユリ(園芸品)(ユリ科)の葯の縦裂開。葯の両縁に下の方から縦の裂け目ができる。

縦裂は上の方へと広がっていき、両縁が先端まで裂ける。


裂け目はさらに広がってほとんど水平に開き、花粉がある葯の内部が葯の表面のほとんどを占めるようになる。
ツツジやナスの仲間のように葯の端の孔から花粉が出るもの(孔開)、クスノキ科のように、葯壁の何ヶ所かが花粉とともにめくれ上がるもの(弁開)もある。


ナス(ナス科)の花は下向きに開き、5つの黄色い葯の先端に2つずつ花粉が出る孔がある
 サツキ(ツツジ科)の葯と柱頭
サツキ(ツツジ科)の葯と柱頭
 左: ゲッケイジュ(クスノキ科)の雄花。葯の向軸側が左右2ヶ所まくれ上がり、花粉を露出させる。
左: ゲッケイジュ(クスノキ科)の雄花。葯の向軸側が左右2ヶ所まくれ上がり、花粉を露出させる。
下: クスノキ(クスノキ科)の葯(SEM画像)。向軸側が4ヶ所まくれ上がる。

葯から露出した花粉は、
-
花粉媒介者(ポリネーター)[pollen vector; pollinator]によって花から運び出され(送粉または花粉送出[pollen export]
-
運ばれて(花粉運搬[pollen transfer])
-
他の花や同じ花の柱頭に付着する(受粉または花粉受取[pollen import])
花粉表出から花粉受取までの一連のプロセスを送受粉(ポリネーション)[pollination]という。ただし、「送粉」または「受粉」で送受粉全体を指すこともあって、花粉を出す花や花粉媒介者から見るときに送粉、花粉を受け取る花から見るときに受粉が使われるようだ。このテキストでは(特に断らないときは)送受粉のことを単に「送粉」と呼ぶ。
葯から出た花粉がいったん花被や花柱に付着し、そこから花粉媒介者に渡るしくみ(二次的花粉表出(仮訳)[secondary pollen presentation])を持つ花も多い(キク科・キキョウ科など)。→
二次的花粉表出
6-1-3. 有性生殖の基本器官 (2)雌しべ
 雌しべ・花粉管の伸長のようすの模式図。図の雌しべは2つの心皮でできており、子房室や柱頭の枝の数に反映している。
雌しべ・花粉管の伸長のようすの模式図。図の雌しべは2つの心皮でできており、子房室や柱頭の枝の数に反映している。
1―柱頭、2―花柱、3―子房、4―心皮、5―花粉粒、6―花粉管、7―子房室、8―子房壁、9―胚珠。
雌しべを機能で区切ると、先端から「柱頭」「花柱」「子房」の3つの部分に分けられる。雌しべのうち、胚珠[ovule](将来、種子になる粒)が入っている部分を子房[ovary]という。子房の中に胚珠が1個だけ入っているときもあれば、多数の胚珠が並んでいるときもある。
「子房」の「子」は胚珠や種子、「房」は部屋(例えば、「厨房」「暖房機」)の意味で使われている。動物学では"ovary"は卵巣、"ovule"は(小型の)卵を指す。
雌しべを形態的な起源によって単一または複数の「心皮」に分けることもある。7-2. 雌しべと心皮で詳しく説明するが、花被片1枚や雄しべ1本が1枚の葉に相当するように、心皮1つが1枚の葉に相当する。雌しべを構成する心皮の数は、柱頭の枝や裂け目の数、花柱が複数ある場合は花柱の数、子房が複数の子房室を持つ場合は子房室の数、子房の断面が角張っている場合は辺の数などと一致することが多い。

サクラ(カラミザクラ)の雌しべ(左)・柱頭(右上)・子房の縦断面(右下)。
雌しべの先端には花粉が付着するところ(柱頭[stigma])があり、たいていは付着しやすいように、粘り気のある毛が密生していたり、全体が粘液で光ったりしている。
 リュウキュウアサガオ(ヒルガオ科)の柱頭(SEM写真)。塊状に集まった大きな突起に細い突起が密生した複雑な構造をしている。
リュウキュウアサガオ(ヒルガオ科)の柱頭(SEM写真)。塊状に集まった大きな突起に細い突起が密生した複雑な構造をしている。
 セイヨウカラシナ(アブラナ科)の柱頭(SEM写真)。突起が密生したドーム状で、上に一筋のくぼみがある。
セイヨウカラシナ(アブラナ科)の柱頭(SEM写真)。突起が密生したドーム状で、上に一筋のくぼみがある。

タカサゴユリ(ユリ科)の柱頭


ムラサキツユクサ(ツユクサ科)柱頭(半分)の押し潰し像。表面突起と付着花粉が見られる。


ダイコン(アブラナ科)柱頭の押し潰し像
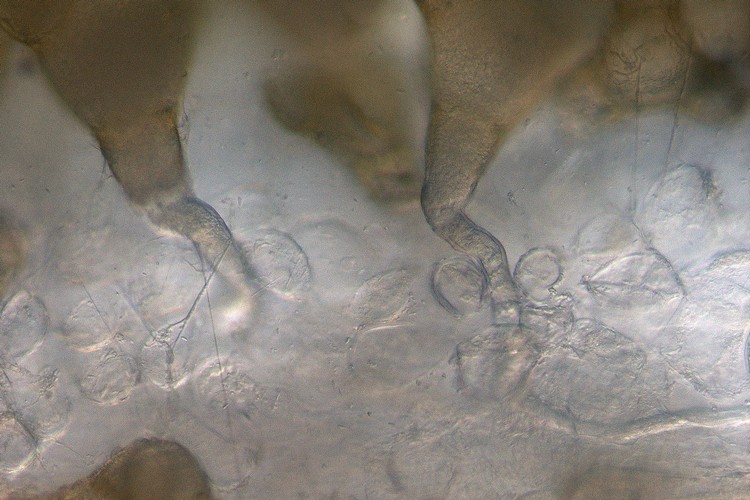
ヒルザキツキミソウ(アカバナ科)柱頭上の発芽花粉と花粉管伸長
受粉滴

 イチョウ(イチョウ科)とヒノキ(ヒノキ科)の胚珠先端に出ている受粉滴
イチョウ(イチョウ科)とヒノキ(ヒノキ科)の胚珠先端に出ている受粉滴
多くの裸子植物では、胚珠の入口(珠孔)から分泌される水滴(受粉滴[pollination drop])が花粉を付着させるはたらきを持つ。付着した花粉は受粉滴ごと胚珠に吸い込まれ、花粉管を出す。
上の定義をきびしく使うと、「柱頭=花粉が付着・発芽する面」ということになるが、花粉の付着・発芽面を含む花柱先端部をまとめて「柱頭」と呼ぶことも多い。




ツクシキケマン(左の2つ)とムラサキケマン(右の2つ)(ともにケシ科|ケマンソウ科)の「柱頭」(それぞれ、左はつぼみ、右は開花後)。半透明の突起部に花粉が付着・発芽する。
ツクシキケマンの柱頭の場合、次の2つの表わし方があるが、図鑑類では2番目のように表わされるのがふつうだ。
-
花柱の先端が弧状に広がり、そのヘリに約10個のイボのような柱頭がついている
-
柱頭は弧状で、へりに約10個の突起(花粉付着部)がある
子房が花の奥の方にあることが多いのに対し、柱頭は外からよく見えるところにあることが多い。だから、たいていは、子房と柱頭は離れている。両者をつなぐ柱状の部分を花柱[style]という。ものすごく長い花柱を持つ花もあれば、わりと短い花柱を持つ花、子房にじかに柱頭がついている(花柱がない)花もある。
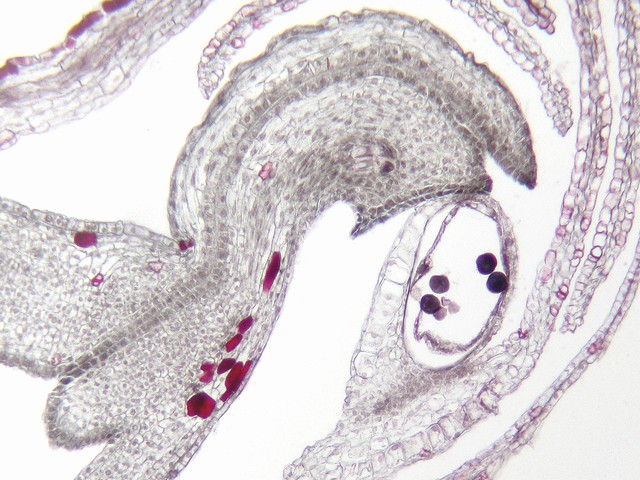
ナガバタチツボスミレ(スミレ科)の花(閉鎖花=開花しないまま結実する花)の雌しべの上部の断面。柱頭から花柱を中心を経て子房室(左下)まで、花粉管の通り道に沿って細胞質が多い(よく染まっている)細胞が続いている。花粉管の発芽と伸長は、これらの細胞に助けられる。
柱頭についた花粉から花粉管[pollen tube]が伸び出して、柱頭から雌しべの中に入る。花粉管は花柱の内部を通って、子房の内部に達し胚珠に入って受精[fertilization]が起こる。

ネズミモチ(モクセイ科)の柱頭。花粉粒には3つのスリット(発芽口)があり、スリットから伸び出した花粉管が柱頭の組織に入っていく。


カラスノエンドウの花粉管。柱頭についた花粉から花粉管が伸び、柱頭の中に入り込む。花柱を通り抜けて子房の中に達した花粉管は胚珠に入る。アニリンブルーで染色し、蛍光顕微鏡で観察した画像。
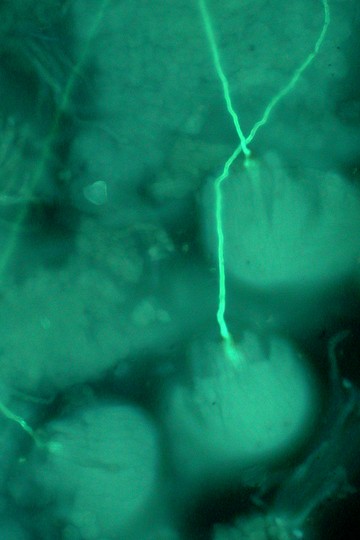

アケビ(アケビ科)の胚珠と花粉管。花粉管の先端は、2枚の種皮に包まれた珠心に入り込む。
6-1-4. 雄しべや雌しべがない花もある

有性生殖器官としての花には、雄花(左上)・雌花(右上)・両性花(下)の3つがある。
両性花と単性花・両性個体と単性個体・集団の雌雄性
雄しべ・雌しべの両方とも機能している花を両性花[bisexual flower; またはhermaphroditic flower]という。一方、雄しべのみが機能している花を雄花[male flower]、雌しべのみが機能している花を雌花[female flower]といい、総称して単性花[unisexual flower]という。
雄花に機能しない雌しべがある、あるいは、雌花に機能しない雄しべがある場合もあり、痕跡的な雌しべ/雄しべ(退化的な雌しべ/雄しべ・仮雌蕊/仮雄蘂)と呼ばれる。このような花を「形態上の両性花で機能上の雄(雌)花」と呼ぶこともあったが、一貫性のある用法とは言い難く、避けた方が良い。
花が両性花のみなら、集団は両性個体のみからなる。被子植物の種の約7割は両性花のみをつけ、残る約3割は雄花と雌花の両方または一方を持つと見積もられている。
一方、花が単性であっても、個体としては、両性である場合も単性である場合もある。前者の代表は「雌雄同株」(より正確には、雌雄異花同株[monoecy])で、雌花と雄花が同じ個体につく(キュウリ・アケビなど)。




キュウリ(ウリ科)は雌雄異花同株で、雌花と雄花がつく
 アケビ(アケビ科)は雌雄異花同株。雄花は数個が一つの花序につき、雌花は単独でつく。雌花には雌しべがたくさんある。アケビの実は、受粉した雌しべが大きくなったもの。
アケビ(アケビ科)は雌雄異花同株。雄花は数個が一つの花序につき、雌花は単独でつく。雌花には雌しべがたくさんある。アケビの実は、受粉した雌しべが大きくなったもの。


後者の場合、雄株(雄花のみをつける個体)か雌株(雌花のみをつける個体)、またはその両方が集団に含まれることになる。集団が雄株と雌株のみからなるとき(マユミなど)、「集団は雌雄異株[dioecy]である」という。


マユミ(ニシキギ科)は雌雄異株で、雄花(退化的な雌しべがある)のみをつける雄個体と雌花(退化的な雄しべがある)のみをつける雌個体に分かれる
両性花のみの種、雌雄異花同株の種、雌雄異株の種だけでなく、単性花と両性花、単性個体と両性個体のさまざまな組み合わせがあり、被子植物の雌と雄の組み合わせ(雌雄性、あるいは性表現ともいう)は非常に多様だ(→植物の雌雄性: タイプ分け)。コケ植物・シダ植物・裸子植物では、有性生殖器官は単性なので、雌雄同株か雌雄異株のいずれかで、コケ植物(配偶体)は雌雄異株が多数派、シダ植物(胞子体)は雌雄同株が圧倒的に多く、裸子植物は約6割が雌雄異株、約4割が雌雄同株だ。
対照的に動物(多細胞動物)では雌雄異体が多数を占める。また、雌雄性は植物に比べて変化に乏しく、雌雄異体の他は、雌雄の生殖器1つずつを持つ(同時的)雌雄同体(カタツムリ・ミミズなど)と条件によって生殖器が雄←雌/雄→雌と変化する異時的雌雄同体(魚類の一部など)の例があるだけだ。
無性花
雄しべ・雌しべのどちらもない(または機能していない)花は無性花(中性花)[asexual flower]という。無性花は、特殊な場合をのぞき、両性花・雌花・雄花の周りを取り巻いて装飾花の役割をする。

シロタエヒマワリ(キク科)。小さい花が集まって密な円盤状の花序(頭状花序)をつくる。花序内側に密集する花(筒状花)は濃黄色~黒褐色の花冠を持ち、両性花で雄性期(1)から雌性期(2)へと変化する。花序の縁にある黄色のへら状花冠を持つ花(舌状花―3)は無性花。 | 
 |
 オオカメノキ(スイカズラ科|レンプクソウ科)。外側の花は花冠が極端に大きいが雄しべ・雌しべを持たない無性花になっており「装飾花」と呼ばれる。中心部には花冠が小さい両性花が集まっている。
オオカメノキ(スイカズラ科|レンプクソウ科)。外側の花は花冠が極端に大きいが雄しべ・雌しべを持たない無性花になっており「装飾花」と呼ばれる。中心部には花冠が小さい両性花が集まっている。
6-1-5. 花被の基本的なはたらき=つぼみの保護と動物の誘引
花被片で構成される花被は雌しべと雄しべの周りを取り巻いている。花被の主な役割は2つに分けられる。
- 保護: 花がまだ咲いていないとき(つぼみのとき)に、花の他の部分を包んで物理的損傷や乾燥、食害を防ぐ
- 誘引: 送粉動物に対して花を目立たせる。
誘引の役割は動物媒花に限られる。風や水で送粉される花の花被は、その必要がないため、地味な色で小さかったり、あるいは、完全に欠落している場合もある。


パーツの数・配置が同じ虫媒花・アマナ(ユリ科)と風媒花・クサイ(イグサ科)の比較
典型的な動物媒花では、外側の花被は保護に専門化して「萼」(花被片は「萼片」)と呼ばれ、内側の花被は誘引に専門化して「花冠」(花被片は「花弁」)と呼ばれる(異花被花[heterochlamydeous flower])。しかし、花冠と萼が分業していないものもあり、ユリ・タブノキのように内外の花被の両方が保護・誘引の両方を担うもの(同花被花[homochlamydeous flower])、ソバのように花被が一重のみで保護・誘引の両方を担うもの(単花被花[monochlamydeous flower])、センリョウのように花被がないもの(無花被花[monochlamydeous flower])もある。
|
異花被花 |
同花被花 |
単花被花 |
無花被花 |
| 外花被 |
萼(保護器官) |
保護+誘引 |
保護+誘引 |
なし |
| 内花被 |
花冠(誘引器官) |
保護+誘引 |


同花被花・ユリ(ユリ科)。つぼみの段階では緑色で堅い内外の花被片が、開花の過程で成長・着色する。


左―単花被花・ソバ(タデ科); 右―無花被花・センリョウ(センリョウ科)
 離弁花(上)と合弁花(下)の模式図。右は発生初期の模式図で、5つの花弁原基(●)が独立したまま発生が進めば離弁花冠、原基どうしの間をつなぐように分裂組織(灰色の部分)ができると合弁花冠となる。
離弁花(上)と合弁花(下)の模式図。右は発生初期の模式図で、5つの花弁原基(●)が独立したまま発生が進めば離弁花冠、原基どうしの間をつなぐように分裂組織(灰色の部分)ができると合弁花冠となる。
異花被花において、花弁が互いにつながり合って一体化している花冠を「合弁花冠」(合弁花冠をもつ花は「合弁花」)、花弁がつながっていない花冠を「離弁花冠」(離弁花冠をもつ花は「離弁花」)という。コヒルガオのように花弁全体がほぼ一体化しているものもあれば、ツバキやエゴノキのように基部のみでつながっているものもある。


ツツジ類は典型的な合弁花(左)だが、園芸品種には離弁花をもつものがあり「采咲き」と呼ばれる(右)。おそらく、離弁←→合弁の移行が比較的単純な遺伝的変化でもたらされることを示している。

離弁花の例: カラミザクラ(バラ科)。花弁は一枚ずつばらばらに「散る」。



合弁花の例(1): 左―コヒルガオ(ヒルガオ科)。開花後の花は萎んでから脱落する。右―ヤブツバキ(ツバキ科)。花弁と雄しべが、丸ごとぽとりと落ちる。





合弁花の例(2): 上―エゴノキ(エゴノキ科)。花弁と雄しべが、丸ごとぽとりと落ちる。下―ローゼル(アオイ科フヨウ属)。花弁と雄しべは萎んでから脱落する。
離弁花類と合弁花類
2000年代初めまで図鑑を含む植物関係の書籍や学校教育で採用されていた「エングラー分類系[Engler system]」、正確には「新エングラー分類系」と呼ばれる分類系*では、被子植物を双子葉植物と単子葉植物に二分し、双子葉植物を離弁花類(古生花被亜綱)と合弁花類(後生花被亜綱)に二分する。
離弁花類/合弁花類では、様々な特徴において原始的な特徴を多く留めると想定されたグループが離弁花類、進化的な特徴を多く持つと想定されたグループが合弁花類に分類された。離弁花・合弁花の違いで単純に分割された訳ではないため、離弁花類に含まれ合弁花をもつ種(アオイ科フヨウ属やウリ科のキュウリ・カボチャ等)や、逆に合弁花類に含まれ離弁花をもつ種(ツツジ科ホツツジ等)が多数ある。
*)ドイツの植物学者エングラー(Adolf Engler; 1844―1930)年10月10日)が提唱し、後の植物学者による修正を経た分類系
保護または誘引、あるいはその両方の役割を花被以外の器官が代行する場合がある。
 ドクダミ(ドクダミ科)。「白い4枚の花びら」のように見えるのは花びらそっくりになった葉で、花序(黄色いところ)を拡大すると、雌しべと雄しべだけの花(無花被花)がぎっしりと集まっている。
ドクダミ(ドクダミ科)。「白い4枚の花びら」のように見えるのは花びらそっくりになった葉で、花序(黄色いところ)を拡大すると、雌しべと雄しべだけの花(無花被花)がぎっしりと集まっている。

ブーゲンビリア(園芸品)(オシロイバナ科)。3枚の色づいた葉が枝先に集まり、それぞれの葉の腋に1つずつ、筒状の単花被花がつく。
6-1-6. 花の部品配置と花式図
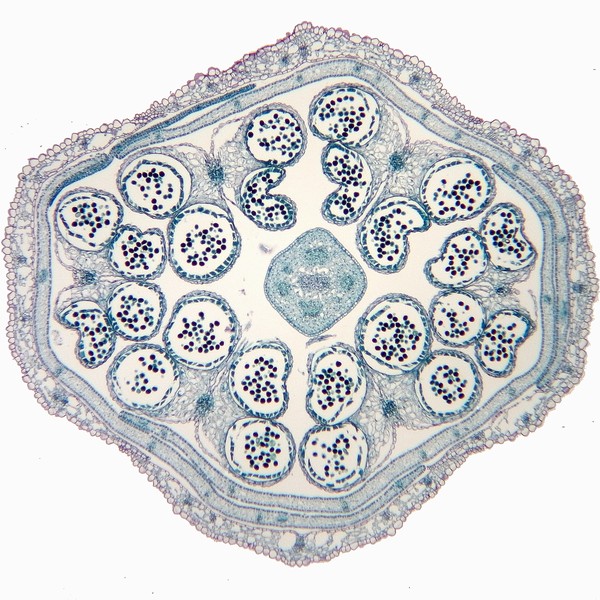

左―セイヨウカラシナ(アブラナ科)・右―ユリ園芸品(ユリ科)のつぼみの横断面。つぼみの方が花の部品配置が分かりやすいことが多い。
花を構成する部品/パーツ[floral parts](花被片と雄しべ・雌しべなど)の数や配置はさまざまで、植物の系統関係と機能の両方を反映する。

分解図に基づく配置図。輪を同心円として描き、部品を配置している。雌しべを子房横断面に置き換えると後出の花式図となる。
上で述べたとおり、外側から萼片・花弁・雄しべ・雌しべの順にらせんを描くように、あるいは同心円(輪 りん[whorl]という)上に並ぶ。同種の部品(例えば花弁)が複数の輪に分かれて並ぶこともあるので、輪の数にも違いがある。1つの輪にある部品の数、また、隣り合った輪にある部品との位置関係も、配置を特徴つける。
数と配置を、花の横断面を見たかたちで模式的に表した図を花式図[floral diagram]という。花式図は写実的である必要はなく、また、配置や数に対する解釈を含んでもよい。例えば、同じ輪にある部品を点線の円の上に並べたり、本来あるべき部品がないと考えられるときは「×」で示すこともある。
また、数と配置を、記号式で表わしたものを花式[floral formula, 複 floral formulae]という。
花式図・花式は、部品の数と位置関係を抽出することで、さまざまな花の特徴を把握しやすく、異なる花どうしを比較しやすくする。両者とも、書き方に決められたルールはないが、19世紀ドイツの植物学者Eichlerの古典的な著作"Blütendiagramme"(花式図)が一つの標準になっている。
-
Eichler AW. 1875. Blütendiagramme, Volume I. Verlag von Wilhelm Engelmann. link
-
Eichler AW. 1878. Blütendiagramme, Volume II. Verlag von Wilhelm Engelmann. link
-
Ronse De Craene LP. 2010. Floral Diagrams: An Aid to Understanding Flower Morphology and Evolution. Cambridge University Press.
アブラナ科
 |
 |
 |
 |
 |
 |
 |
 |
|
セイヨウアブラナ(アブラナ科)の花と花式図。雌しべ1個・雄しべ6本(短い雄しべ2本+長い雄しべ4本)・花弁4枚・萼片4枚がある。基部には、緑色の蜜腺が4つあり、蜜の滴を分泌する。 |
セイヨウアブラナの花式図。アブラナ科の多くの種は、同じ花式図で表現できる。上段は部品の配置のみを表した花式図、下段は配置に関する解釈を含んだ花式図。
1―萼片・2―花弁・3―雄しべ(短)・(3)―欠失した雄しべ・4―雄しべ(長)・5―蜜腺・6~9―雌しべ(6―子房壁・7―子房室・8―胚珠・9―隔壁)。
雌しべ断面が実際から90°回転している間違った花式図を掲載していました。岡本素治さんに指摘いただき、訂正しました(2025年4月)。
セイヨウアブラナの花(そして、多くのアブラナ科の花)は、次のように、5つの輪があり、各輪の部品の基本数は4、と解釈される。
| 輪 |
1 |
2 |
3 |
4 |
5 |
| 部品名 |
萼片 |
花弁 |
雄しべ |
心皮 |
| 数 |
4 |
4 |
2 (2個が欠失) |
4 |
2 |
花式: K4 C4 A2+4 G(2)
K―萼片、C―花弁、A―雄しべ、G―雌しべ。Gの下線は上位子房を、(2)は2つの部品が融合して1体となっていることを示す。
ユリ



 ユリ属の花と花式図。同じ花式図は多数の単子葉植物にあてはまる。
ユリ属の花と花式図。同じ花式図は多数の単子葉植物にあてはまる。
| 輪 |
1 |
2 |
3 |
4 |
5 |
| 部品名 |
花被片 |
雄しべ |
心皮 |
| 外花被片 |
内花被片 |
| 数 |
3 |
3 |
3 |
3 |
3 |
花式: P3+3 A3+3 G(3)
P―花被片、A―雄しべ、G―雌しべ。Gの下線は上位子房を、(3)は3つの部品が融合して1体となっていることを示す。
タブノキ

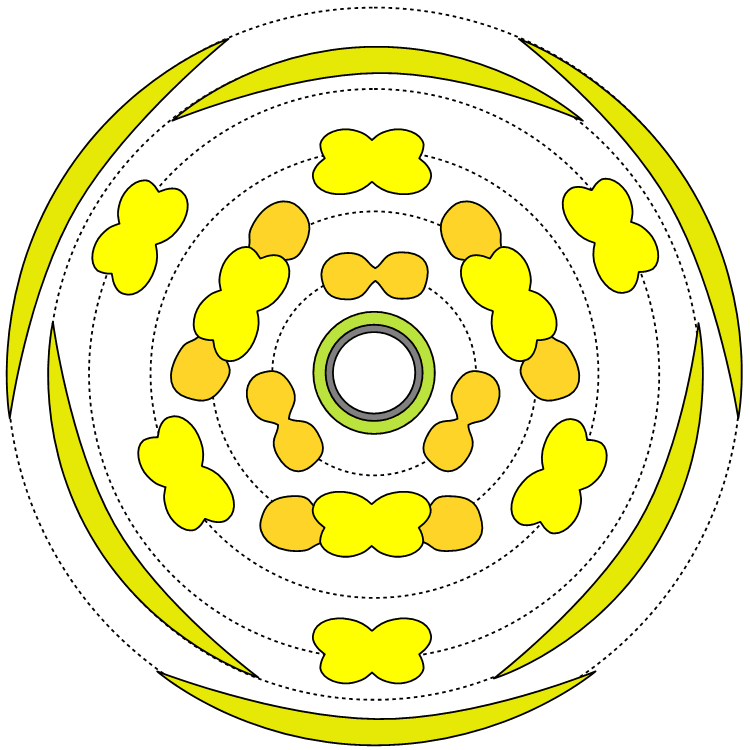
タブノキ(クスノキ科)の花と花式図
| 輪 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
| 部品名 |
花被片 |
雄しべ |
仮雄しべ
(蜜腺) |
心皮 |
| 外花被片 |
内花被片 |
| 数 |
3 |
3 |
3 |
3 |
3 |
3 |
1 |
花式: P3+3 A3+3+3+3° G1
P―花被片、A―雄しべ、G―雌しべ。Aの最後の3°は最内輪が仮雄しべ(雄しべ由来だが花粉を出さない=雄しべ本来の機能がない器官)を、Gの下線は上位子房を示す。
配置以上の情報を花式図に盛り込むこともできる。部品どうしがつながっている状態(合着)は、花式図では、部品どうしを実線でつなぐことで表現する。
合着には、同じ種類の部品どうしのつながり(花弁どうし、花糸どうし、など)と異なる部品どうしのつながり(花弁と雄しべ、など)がある。多くの場合は、部品の形成過程で原基と原基の間が成長することでつながるようになるが、形成後の部品の縁どうしがかみ合うようにしてつながる場合もある。
ナス(ナス科)の花

 ナス科の多くの種にあてはまる花式図。1―萼片、2―花弁、3―雄しべ、4~7―雌しべ(4―子房壁、5―胎座、6―子房室、7―胚珠)。隣り合う花弁どうしを結ぶ実線(a)によって花弁どうしがつながっていること(合弁)が、雄しべと花弁を結ぶ実線(b)によって雄しべが花弁の途中についていることが表現されている。
ナス科の多くの種にあてはまる花式図。1―萼片、2―花弁、3―雄しべ、4~7―雌しべ(4―子房壁、5―胎座、6―子房室、7―胚珠)。隣り合う花弁どうしを結ぶ実線(a)によって花弁どうしがつながっていること(合弁)が、雄しべと花弁を結ぶ実線(b)によって雄しべが花弁の途中についていることが表現されている。

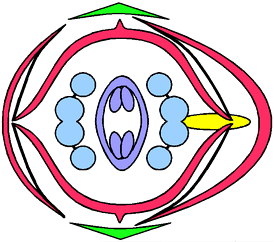 キケマン属(ケマンソウ科)の花式図の例(写真はムラサキケマン)。キケマン属は左右相称のやや複雑な花を持つ(→ムラサキケマン)。実線でつなぐことで花弁どうし、雄しべどうしの合着を、花弁をふくらませて距を表わしている。
キケマン属(ケマンソウ科)の花式図の例(写真はムラサキケマン)。キケマン属は左右相称のやや複雑な花を持つ(→ムラサキケマン)。実線でつなぐことで花弁どうし、雄しべどうしの合着を、花弁をふくらませて距を表わしている。
花式でも、記号を追加することで情報を追加できる(例: Ronse De Craene 2010: 39)が、かなり複雑になる。
基本数と互生則
上の5例では、次の2つが成り立っていた。
-
心皮をのぞいては、各輪の部品数は一定で、「基本数」を決めることができる
-
隣り合う輪では、部品は互い違いに配置する(互生則)
各輪の部品の基本数がXであるとき、「X数性の花」という。アブラナは4数性[tetramerous]、ユリ・タブノキは3数性[trimerous]、ナスは5数性[pentamerous]、キケマン属は2数性[dimerous]の花を持つ。
この2つが成り立つ場合が多いことは確かだが、例外も少なくない。
アオカズラは、萼片・花弁・雄しべがいずれも同じ位置にある(対生している)。
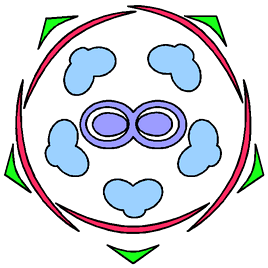
アオカズラ(アワブキ科)の花式図
タデ類・ミゾソバ・イタドリなどでは、花被片と外輪の雄しべは5個ずつ、内輪の雄しべは3個となる。


タデ科に多く見られる花式図(写真はママコノシリヌグイ)
一つの花序で基本数が違う花が混じる例もある。

ヘンルーダ(ミカン科)の花序。中心の頂花は5数性、他の花は4数性。
テキスト目次に戻る
ホームに戻る









